Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada.
La única característica de la multiplicación viral consiste en que, poco después de su interacción con una célula huésped, el virión infectante se desintegra y pierde la infecciosidad que se puede medir. Esta fase del ciclo celular se llama periodo de eclipse; su duración varía dependiendo del virus y de la célula huésped en particular, y termina con la formación de las primeras partículas virales descendientes infecciosas. El periodo de eclipse es, en realidad, de intensa actividad sintética, puesto que la célula se redirige hacia la satisfacción de las necesidades del "pirata" viral. En algunos casos, tan pronto como el ácido nucleico viral penetra a la célula huésped, el metabolismo celular es recanalizado exclusivamente hacia la síntesis de nuevas partículas virales. En otros casos, los procesos metabólicos de la célula huésped no se alteran significativamente, aunque la célula sintetiza proteínas virales y ácidos nucleicos.
Los virus han desarrollado diversas estrategias para lograr la multiplicación en las células huéspedes parasitadas por ellos. Aunque los detalles varían de un grupo a otro, el patrón general de los ciclos de replicación es semejante.
La primera etapa de la infección viral es la interacción de un virión con un sitio receptor específica de la superficie celular. Las moléculas receptoras difieren para los distintos virus, y son proteínas en algunos casos (por ejemplo, picornavirus) y oligosacáridos en otros (por ejemplo, ortovirus y paramixovirus). La presencia o la ausencia de receptores desempeña una función determinante de primer orden en el tropismo celular y la patogenia viral; por ejemplo, el poliovirus es capaz de fijarse sólo en las células del sistema nervioso central y las vías digestivas de los primates. Se cree que la fijación en receptores refleja homologías configurativas fortuitas entre la estructura de la superficie del virión y un componente de la superficie celular. Por ejemplo, el virus de la inmunodeficiencia humana (HIV) se une al receptor CD4 en las células del sistema inmunitario y se cree que el virus de la rabia interactúa con los receptores de acetilcolina y que el virus de Epstein-Barr reconoce al receptor para el tercer componente del complemento en las células B. Es probable que cada célula susceptible posea por lo menos 100,000 sitios receptores para un virus dado.
Después de la fijación, la partícula viral entra en la célula. Esta etapa se conoce como penetración, viropexia o fagocitosis. En algunos sistemas, se realiza por endocitosis mediada por receptores, con captación de las partículas virales ingeridas dentro de endosomas. Están menos claros los detalles de la penetración en otros sistemas. Ocurre perdida de la cubierta de manera concomitante con la penetración o poco después de la misma. La pérdida de la cubierta, o descubrimiento, es la separación física del ácido nucleico viral (o, en algunos casos, de las nucleocápsides inteARNs) a partir de los componentes estructurales externos del virión. En este momento se pierde la infecciosidad del virus original.
Los virus son los únicos agentes infecciosos en los que la disolución del agente infectante es una etapa obligatoria de la vía de la replicación.
Poco después de quedar descubierto el genoma viral, se inicia la fase sintética del ciclo de replicación del virus. El aspecto esencial de la replicación viral consiste en que deben transcribirse los ARNm específicos del ácido nucleico viral para que se exprese y duplique con éxito la información genética. Una vez que se logra esta etapa, los virus recurren a los componentes de la célula para traducir el ARNm. Diversas clases de virus emplean vías diferentes para sintetizar los ARNm según la estructura del ácido nucleico viral. Algunos virus (por ejemplo, rhabdovirus, ortomixovirus y paramixovirus) poseen ARN polimerasas que sintetizan a los ARNm. Los virus ARN de este tipo se llaman virus de tira negativa (de sentido negativo), puesto que su genoma de ARN de una sola tira es complementario del ARNm, el cual se designa por acuerdo general como de tira positiva (o de sentido positivo).
Durante la replicación viral se sintetizan todas las macromoléculas especificadas del virus en una secuencia muy organizada. En algunas infecciones por virus, notablemente las producidas por virus de doble tira que contienen ADN, se sintetizan proteínas virales tempranas poco después de la infección y aparecen proteínas tardías sólo durante la fase tardía del proceso infeccioso, una vez que ha ocurrido la síntesis del ADN viral. Pueden activarse o no genes tempranos cuando se elaboran los productos tardíos. En contraste, se expresa al mismo tiempo la mayor parte de la información genética de los virus que contienen ARN o toda ella. Además de estos controles temporales, existen también controles cuantitativos puesto que no se elaboran todas las proteínas virales en las mismas cantidades. Las proteínas específicas del virus pueden regular la extensión de la transcripción del genoma o la traducción del ARNm viral.
Se encuentra la variación más amplia en las estrategias de la expresión genética entre los virus que contienen ARN. Algunos viriones poseen polimerasas (ortomixovirus, reovirus); otros sistemas emplean mensajes subgenómicos generados a veces por empalme (ortomixovirus, retrovirus); por último, algunos virus sintetizan grandes precursores poliproteínicos que se someten a procesamiento y segmentación para generar los productos génicos finales (picornavirus, retrovirus).
La extensión a la que las enzimas específicas del virus participan en estos procesos varía de un grupo a otro. En general, los virus de mayor tamaño (herpesvirus, poxvirus) son más independientes de las funciones celulares que los virus de menor tamaño. Esta es una razón de que los virus de mayor tamaño sean más sensibles a la quimioterapia antiviral, porque se dispone de más procesos específicos de los virus como blancos de la acción farmacológica.
Los sitios intracelulares en los que ocurren los diferentes acontecimientos de la replicación viral, varían de un grupo a otro. Es posible plantear algunas generalizaciones. La proteína viral se sintetiza en el citoplasma, sobre los polirribosomas compuestos por ARNm específico del virus y ribosomas de la célula huésped. El ADN viral suele replicarse en el núcleo. El ARN genómico viral se duplica generalmente en el citoplasma celular, aunque hay excepciones.
1. ADHESION El primer paso en la infección de una célula es la adhesión o fijación a la superficie celular. Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula. Las células que carecen de los receptores apropiados no son susceptibles al virus.
2. PENETRACIÓN Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus.
Virus envueltos
(A) Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular.
(B) Entada vía endosomas en la superficie celular.
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
Virus no envueltos o desnudos Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
3. EMSAMBLAJE Y MADURACION Ensamblaje de los componentes de los virus para formar nuevas partículas víricas (proceso también llamado maduración).
Los virus desnudos realizan su ensamblaje y maduración en forma intracelular y adquieren capacidad infectante dentro de la célula. Para su salida requieren de la desintegración celular. Los virus envueltos en el último paso al ensamblaje el virion se une con la membrana celular a la salida de la célula, mediante proteínas que se insertan en la superficie interna y exterior de la membrana. El virion naciente brota atraves de la membrana. El ensamblaje y maduración con la expulsión atraves de la superficie celular proporcionan un mecanismo más eficaz de salida, ya que no depende de la desintegración de la célula infectada.
4. SALIDA Liberación de los nuevos virus de la célula hospedadora, que ya pueden infectar otras células. Pueden producirse con o sin rotura de la célula, dependiendo de los virus. Los virus desnudos salen por lisis celular lo contrario de los envueltos que salen por gemación.
paula ortiz 88402 1. Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada
88402 La única característica de la multiplicación viral consiste en que, poco después de su interacción con una célula huésped, el virión infectante se desintegra y pierde la infecciosidad que se puede medir. Esta fase del ciclo celular se llama periodo de eclipse; su duración varía dependiendo del virus y de la célula huésped en particular, y termina con la formación de las primeras partículas virales descendientes infecciosas. El periodo de eclipse es, en realidad, de intensa actividad sintética, puesto que la célula se redirige hacia la satisfacción de las necesidades del "pirata" viral. En algunos casos, tan pronto como el ácido nucleico viral penetra a la célula huésped, el metabolismo celular es recanalizado exclusivamente hacia la síntesis de nuevas partículas virales. En otros casos, los procesos metabólicos de la célula huésped no se alteran significativamente, aunque la célula sintetiza proteínas virales y ácidos nucleicos.
88402 Los virus han desarrollado diversas estrategias para lograr la multiplicación en las células huéspedes parasitadas por ellos. Aunque los detalles varían de un grupo a otro, el patrón general de los ciclos de replicación es semejante. Virus no envueltos o desnudos Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
88402 4-La primera etapa de la infección viral es la interacción de un virión con un sitio receptor específica de la superficie celular. Las moléculas receptoras difieren para los distintos virus, y son proteínas en algunos casos (por ejemplo, picornavirus) y oligosacáridos en otros (por ejemplo, ortovirus y paramixovirus). La presencia o la ausencia de receptores desempeña una función determinante de primer orden en el tropismo celular y la patogenia viral; por ejemplo, el poliovirus es capaz de fijarse sólo en las células del sistema nervioso central y las vías digestivas de los primates. Se cree que la fijación en receptores refleja homologías configurativas fortuitas entre la estructura de la superficie del virión y un componente de la superficie celular. Por ejemplo, el virus de la inmunodeficiencia humana (HIV) se une al receptor CD4 en las células del sistema inmunitario y se cree que el virus de la rabia interactúa con los receptores de acetilcolina y que el virus de Epstein-Barr reconoce al receptor para el tercer componente del complemento en las células B. Es probable que cada célula susceptible posea por lo menos 100,000 sitios receptores para un virus dado
88402 5-mediante Después de la fijación, la partícula viral entra en la célula. Esta etapa se conoce como penetración, viropexia o fagocitosis. En algunos sistemas, se realiza por endocitosis mediada por receptores, con captación de las partículas virales ingeridas dentro de endosomas. Están menos claros los detalles de la penetración en otros sistemas. Ocurre perdida de la cubierta de manera concomitante con la penetración o poco después de la misma. La pérdida de la cubierta, o descubrimiento, es la separación física del ácido nucleico viral (o, en algunos casos, de las nucleocápsides inteARNs) a partir de los componentes estructurales externos del virión. En este momento se pierde la infecciosidad del virus original. Los virus son los únicos agentes infecciosos en los que la disolución del agente infectante es una etapa obligatoria de la vía de la replicación
88402 6-durante Poco después de quedar descubierto el genoma viral, se inicia la fase sintética del ciclo de replicación del virus. El aspecto esencial de la replicación viral consiste en que deben transcribirse los ARNm específicos del ácido nucleico viral para que se exprese y duplique con éxito la información genética. Una vez que se logra esta etapa, los virus recurren a los componentes de la célula para traducir el ARNm. Diversas clases de virus emplean vías diferentes para sintetizar los ARNm según la estructura del ácido nucleico viral. Algunos virus (por ejemplo, rhabdovirus, ortomixovirus y paramixovirus) poseen ARN polimerasas que sintetizan a los ARNm. Los virus ARN de este tipo se llaman virus de tira negativa (de sentido negativo), puesto que su genoma de ARN de una sola tira es complementario del ARNm, el cual se designa por acuerdo general como de tira positiva (o de sentido positivo).
Durante la replicación viral se sintetizan todas las macromoléculas especificadas del virus en una secuencia muy organizada. En algunas infecciones por virus, notablemente las producidas por virus de doble tira que contienen ADN, se sintetizan proteínas virales tempranas poco después de la infección y aparecen proteínas tardías sólo durante la fase tardía del proceso infeccioso, una vez que ha ocurrido la síntesis del ADN viral. Pueden activarse o no genes tempranos cuando se elaboran los productos tardíos. En contraste, se expresa al mismo tiempo la mayor parte de la información genética de los virus que contienen ARN o toda ella. Además de estos controles temporales, existen también controles cuantitativos puesto que no se elaboran todas las proteínas virales en las mismas cantidades. Las proteínas específicas del virus pueden regular la extensión de la transcripción del genoma o la traducción del ARNm viral.
88402 7-todo Se encuentra la variación más amplia en las estrategias de la expresión genética entre los virus que contienen ARN. Algunos viriones poseen polimerasas (ortomixovirus, reovirus); otros sistemas emplean mensajes subgenómicos generados a veces por empalme (ortomixovirus, retrovirus); por último, algunos virus sintetizan grandes precursores poliproteínicos que se someten a procesamiento y segmentación para generar los productos génicos finales (picornavirus, retrovirus).
La extensión a la que las enzimas específicas del virus participan en estos procesos varía de un grupo a otro. En general, los virus de mayor tamaño (herpesvirus, poxvirus) son más independientes de las funciones celulares que los virus de menor tamaño. Esta es una razón de que los virus de mayor tamaño sean más sensibles a la quimioterapia antiviral, porque se dispone de más procesos específicos de los virus como blancos de la acción farmacológica.
Los sitios intracelulares en los que ocurren los diferentes acontecimientos de la replicación viral, varían de un grupo a otro. Es posible plantear algunas generalizaciones. La proteína viral se sintetiza en el citoplasma, sobre los polirribosomas compuestos por ARNm específico del virus y ribosomas de la célula huésped. El ADN viral suele replicarse en el núcleo. El ARN genómico viral se duplica generalmente en el citoplasma celular, aunque hay excepciones
1- El primer paso en la infección viral es la absorción a la membrana de la célula susceptible por medio de la adhesión de ligandos virales (proteínas de la capside o glucoproteínas de las espículas, por ejemplo a receptores superficiales de la célula). La distribución en el cuerpo del hospedador de ciertos tipos de receptores celulares explica el tropismo tisular y de hospedador de los virus. Los tejidos y células que carecen de receptores específicos de virus determinados no son infectados por dichos viriones. La naturaleza de los receptores es variable y ciertos virus pueden tener más de un tipo de receptor. 2- Los virus complejos producen una rotura en la membrana celular del hospedador en uno de los puntos de anclaje, gracias a la presencia de algunas moléculas de enzimas hidrolíticas entre lasproteínas de la cápsida. A través de la rotura, el tubo central inyecta el ADN vírico, quedando la cápside vacía en el exterior de la célula diana y el ácido nucleico libre en el citoplasma. La presencia de cápsidas en la superficie celular es un buen indicio de que ha sufrido una infección vírica.
Otros virus sin envoltura lipídica se introducen en la célula con cápsida y todo, lo cual puede realizarse de dos maneras: • Por penetración directa: después de la fijación, el virus abre una brecha en la membrana y se introduce en el citoplasma. • Por endocitosis: la membrana forma una invaginación en torno al virus, llegando a formar una vesícula que penetra en la célula. Formada la vesícula, el virus abre una brecha en la membrana de la misma con ayuda de algunas enzimas hidrolíticas que él mismo transporta, penetrando así en el citoplasma. Los virus con envoltura lipídica burlan la barrera de la membrana celular porque su cubierta lipídica se funde con la membrana, ya que son de la misma naturaleza. Esta fusión de membranas puede realizarse en dos lugares distintos: • Fusión en la superficie celular: de manera que el virión penetra directamente en el citoplasma. • Fusión con un lisosoma: se forma una vesícula por endocitosis, a la que se une un lisosoma para digerir la partícula introducida; entonces, la cubierta lipídica del virus se funde con la membrana del lisosoma y el virión escapa hacia el citoplasma.
3- Según la duración de la fase de eclipse, se suelen distinguir dos modalidades de ciclo infeccioso de un virus: • Ciclo ordinario: el ácido nucleico vírico procede inmediatamente a la transcripción de su mensaje genético en los ARN necesarios para su multiplicación, y prosigue rápidamente el ciclo vital. Este tipo de ciclo es el más extendido en la naturaleza. • Ciclo lisogénico: fue descubierto por André Lwoff en bacteriófagos. El ADN vírico se cierra por sus extremos generando un ADN circular. Este ADN se inserta en el ADN bacteriano en un lugar específico en el que la secuencia de nucleótidos bacterianos es semejante alguna región del ADN vírico.
Algunos virus que infectan células animales siguen también el ciclo lisogénico, como los papilomavirus de las verrugas y algunos retrovirus que producen algunos tipos de cáncer. En el caso de los retrovirus, conviene recordar que el ácido nucleico es ARN monocatenario, por lo que la transcriptasa inversa ha de copiar el genoma vírico en forma de ADN antes de que pueda insertarse en el ADN celular. 4- La multiplicación del virus consta de la replicación de su material genético, de la transcripción de su mensaje en una molécula de ARN y de la traducción del mensaje para producir proteínasvíricas, tanto las que formarán parte de la cápsida como las proteínas enzimáticas necesarias para el ensamblaje de las piezas del virión y para algunas de las funciones anteriores. Losribosomas y la mayor parte de las enzimas que los ácidos nucleicos víricos utilizan en estos procesos son los de la célula infectada.
Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada.
La única característica de la multiplicación viral consiste en que, poco después de su interacción con una célula huésped, el virión infectante se desintegra y pierde la infecciosidad que se puede medir. Esta fase del ciclo celular se llama periodo de eclipse; su duración varía dependiendo del virus y de la célula huésped en particular, y termina con la formación de las primeras partículas virales descendientes infecciosas. El periodo de eclipse es, en realidad, de intensa actividad sintética, puesto que la célula se redirige hacia la satisfacción de las necesidades del "pirata" viral. En algunos casos, tan pronto como el ácido nucleico viral penetra a la célula huésped, el metabolismo celular es recanalizado exclusivamente hacia la síntesis de nuevas partículas virales. En otros casos, los procesos metabólicos de la célula huésped no se alteran significativamente, aunque la célula sintetiza proteínas virales y ácidos nucleicos.
Los virus han desarrollado diversas estrategias para lograr la multiplicación en las células huéspedes parasitadas por ellos. Aunque los detalles varían de un grupo a otro, el patrón general de los ciclos de replicación es semejante.
La primera etapa de la infección viral es la interacción de un virión con un sitio receptor específica de la superficie celular. Las moléculas receptoras difieren para los distintos virus, y son proteínas en algunos casos (por ejemplo, picornavirus) y oligosacáridos en otros (por ejemplo, ortovirus y paramixovirus). La presencia o la ausencia de receptores desempeña una función determinante de primer orden en el tropismo celular y la patogenia viral; por ejemplo, el poliovirus es capaz de fijarse sólo en las células del sistema nervioso central y las vías digestivas de los primates. Se cree que la fijación en receptores refleja homologías configurativas fortuitas entre la estructura de la superficie del virión y un componente de la superficie celular. Por ejemplo, el virus de la inmunodeficiencia humana (HIV) se une al receptor CD4 en las células del sistema inmunitario y se cree que el virus de la rabia interactúa con los receptores de acetilcolina y que el virus de Epstein-Barr reconoce al receptor para el tercer componente del complemento en las células B. Es probable que cada célula susceptible posea por lo menos 100,000 sitios receptores para un virus dado.
Después de la fijación, la partícula viral entra en la célula. Esta etapa se conoce como penetración, viropexia o fagocitosis. En algunos sistemas, se realiza por endocitosis mediada por receptores, con captación de las partículas virales ingeridas dentro de endosomas. Están menos claros los detalles de la penetración en otros sistemas. Ocurre perdida de la cubierta de manera concomitante con la penetración o poco después de la misma. La pérdida de la cubierta, o descubrimiento, es la separación física del ácido nucleico viral (o, en algunos casos, de las nucleocápsides inteARNs) a partir de los componentes estructurales externos del virión. En este momento se pierde la infecciosidad del virus original.
Los virus son los únicos agentes infecciosos en los que la disolución del agente infectante es una etapa obligatoria de la vía de la replicación. Poco después de quedar descubierto el genoma viral, se inicia la fase sintética del ciclo de replicación del virus. El aspecto esencial de la replicación viral consiste en que deben transcribirse los ARNm específicos del ácido nucleico viral para que se exprese y duplique con éxito la información genética. Una vez que se logra esta etapa, los virus recurren a los componentes de la célula para traducir el ARNm. Diversas clases de virus emplean vías diferentes para sintetizar los ARNm según la estructura del ácido nucleico viral. Algunos virus (por ejemplo, rhabdovirus, ortomixovirus y paramixovirus) poseen ARN polimerasas que sintetizan a los ARNm. Los virus ARN de este tipo se llaman virus de tira negativa (de sentido negativo), puesto que su genoma de ARN de una sola tira es complementario del ARNm, el cual se designa por acuerdo general como de tira positiva (o de sentido positivo).
Durante la replicación viral se sintetizan todas las macromoléculas especificadas del virus en una secuencia muy organizada. En algunas infecciones por virus, notablemente las producidas por virus de doble tira que contienen ADN, se sintetizan proteínas virales tempranas poco después de la infección y aparecen proteínas tardías sólo durante la fase tardía del proceso infeccioso, una vez que ha ocurrido la síntesis del ADN viral. Pueden activarse o no genes tempranos cuando se elaboran los productos tardíos. En contraste, se expresa al mismo tiempo la mayor parte de la información genética de los virus que contienen ARN o toda ella. Además de estos controles temporales, existen también controles cuantitativos puesto que no se elaboran todas las proteínas virales en las mismas cantidades. Las proteínas específicas del virus pueden regular la extensión de la transcripción del genoma o la traducción del ARNm viral.
Se encuentra la variación más amplia en las estrategias de la expresión genética entre los virus que contienen ARN. Algunos viriones poseen polimerasas (ortomixovirus, reovirus); otros sistemas emplean mensajes subgenómicos generados a veces por empalme (ortomixovirus, retrovirus); por último, algunos virus sintetizan grandes precursores poliproteínicos que se someten a procesamiento y segmentación para generar los productos génicos finales (picornavirus, retrovirus).
La extensión a la que las enzimas específicas del virus participan en estos procesos varía de un grupo a otro. En general, los virus de mayor tamaño (herpesvirus, poxvirus) son más independientes de las funciones celulares que los virus de menor tamaño. Esta es una razón de que los virus de mayor tamaño sean más sensibles a la quimioterapia antiviral, porque se dispone de más procesos específicos de los virus como blancos de la acción farmacológica.
Los sitios intracelulares en los que ocurren los diferentes acontecimientos de la replicación viral, varían de un grupo a otro. Es posible plantear algunas generalizaciones. La proteína viral se sintetiza en el citoplasma, sobre los polirribosomas compuestos por ARNm específico del virus y ribosomas de la célula huésped. El ADN viral suele replicarse en el núcleo. El ARN genómico viral se duplica generalmente en el citoplasma celular, aunque hay excepciones.
Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada.
La única característica de la multiplicación viral consiste en que, poco después de su interacción con una célula huésped, el virión infectante se desintegra y pierde la infecciosidad que se puede medir. Esta fase del ciclo celular se llama periodo de eclipse; su duración varía dependiendo del virus y de la célula huésped en particular, y termina con la formación de las primeras partículas virales descendientes infecciosas. El periodo de eclipse es, en realidad, de intensa actividad sintética, puesto que la célula se redirige hacia la satisfacción de las necesidades del "pirata" viral. En algunos casos, tan pronto como el ácido nucleico viral penetra a la célula huésped, el metabolismo celular es recanalizado exclusivamente hacia la síntesis de nuevas partículas virales. En otros casos, los procesos metabólicos de la célula huésped no se alteran significativamente, aunque la célula sintetiza proteínas virales y ácidos nucleicos. mat:89206
Los virus han desarrollado diversas estrategias para lograr la multiplicación en las células huéspedes parasitadas por ellos. Aunque los detalles varían de un grupo a otro, el patrón general de los ciclos de replicación es semejante
La primera etapa de la infección viral es la interacción de un virión con un sitio receptor específica de la superficie celular. Las moléculas receptoras difieren para los distintos virus, y son proteínas en algunos casos (por ejemplo, picornavirus) y oligosacáridos en otros (por ejemplo, ortovirus y paramixovirus). La presencia o la ausencia de receptores desempeña una función determinante de primer orden en el tropismo celular y la patogenia viral; por ejemplo, el poliovirus es capaz de fijarse sólo en las células del sistema nervioso central y las vías digestivas de los primates. Se cree que la fijación en receptores refleja homologías configurativas fortuitas entre la estructura de la superficie del virión y un componente de la superficie celular. Por ejemplo, el virus de la inmunodeficiencia humana (HIV) se une al receptor CD4 en las células del sistema inmunitario y se cree que el virus de la rabia interactúa con los receptores de acetilcolina y que el virus de Epstein-Barr reconoce al receptor para el tercer componente del complemento en las células B. Es probable que cada célula susceptible posea por lo menos 100,000 sitios receptores para un virus dado. mat:89206
Después de la fijación, la partícula viral entra en la célula. Esta etapa se conoce como penetración, viropexia o fagocitosis. En algunos sistemas, se realiza por endocitosis mediada por receptores, con captación de las partículas virales ingeridas dentro de endosomas. Están menos claros los detalles de la penetración en otros sistemas. Ocurre perdida de la cubierta de manera concomitante con la penetración o poco después de la misma. La pérdida de la cubierta, o descubrimiento, es la separación física del ácido nucleico viral (o, en algunos casos, de las nucleocápsides inteARNs) a partir de los componentes estructurales externos del virión. En este momento se pierde la infecciosidad del virus original.
Los virus son los únicos agentes infecciosos en los que la disolución del agente infectante es una etapa obligatoria de la vía de la replicación.
Poco después de quedar descubierto el genoma viral, se inicia la fase sintética del ciclo de replicación del virus. El aspecto esencial de la replicación viral consiste en que deben transcribirse los ARNm específicos del ácido nucleico viral para que se exprese y duplique con éxito la información genética. Una vez que se logra esta etapa, los virus recurren a los componentes de la célula para traducir el ARNm. Diversas clases de virus emplean vías diferentes para sintetizar los ARNm según la estructura del ácido nucleico viral. Algunos virus (por ejemplo, rhabdovirus, ortomixovirus y paramixovirus) poseen ARN polimerasas que sintetizan a los ARNm. Los virus ARN de este tipo se llaman virus de tira negativa (de sentido negativo), puesto que su genoma de ARN de una sola tira es complementario del ARNm, el cual se designa por acuerdo general como de tira positiva (o de sentido positivo).
Durante la replicación viral se sintetizan todas las macromoléculas especificadas del virus en una secuencia muy organizada. En algunas infecciones por virus, notablemente las producidas por virus de doble tira que contienen ADN, se sintetizan proteínas virales tempranas poco después de la infección y aparecen proteínas tardías sólo durante la fase tardía del proceso infeccioso, una vez que ha ocurrido la síntesis del ADN viral. Pueden activarse o no genes tempranos cuando se elaboran los productos tardíos. En contraste, se expresa al mismo tiempo la mayor parte de la información genética de los virus que contienen ARN o toda ella. Además de estos controles temporales, existen también controles cuantitativos puesto que no se elaboran todas las proteínas virales en las mismas cantidades. Las proteínas específicas del virus pueden regular la extensión de la transcripción del genoma o la traducción del ARNm viral. mat:89206
Se encuentra la variación más amplia en las estrategias de la expresión genética entre los virus que contienen ARN. Algunos viriones poseen polimerasas (ortomixovirus, reovirus); otros sistemas emplean mensajes subgenómicos generados a veces por empalme (ortomixovirus, retrovirus); por último, algunos virus sintetizan grandes precursores poliproteínicos que se someten a procesamiento y segmentación para generar los productos génicos finales (picornavirus, retrovirus).
La extensión a la que las enzimas específicas del virus participan en estos procesos varía de un grupo a otro. En general, los virus de mayor tamaño (herpesvirus, poxvirus) son más independientes de las funciones celulares que los virus de menor tamaño. Esta es una razón de que los virus de mayor tamaño sean más sensibles a la quimioterapia antiviral, porque se dispone de más procesos específicos de los virus como blancos de la acción farmacológica.
Los sitios intracelulares en los que ocurren los diferentes acontecimientos de la replicación viral, varían de un grupo a otro. Es posible plantear algunas generalizaciones. La proteína viral se sintetiza en el citoplasma, sobre los polirribosomas compuestos por ARNm específico del virus y ribosomas de la célula huésped. El ADN viral suele replicarse en el núcleo. El ARN genómico viral se duplica generalmente en el citoplasma celular, aunque hay excepciones. mat:89206
Diapositiba1 Etapas de la replicación viral La replicación es el proceso en el cual se copia el ADN. Este proceso es semiconservativo y bidireccional. Funciona igual, tanto en procariontes como en eucarionte, salvo algunos cambio, principalmente de las proteínas que participan. Se dieron muchas hipótesis sobre como se dupllicaba el ADN hasta que Watson y Crick propusieron la hipótesis semiconservativa, según la cual, las nuevas moléculas de ADN formadas a partir de otra antigua, tienen una hebra antigua y otra nueva. La replicación ocurre en 3 etapas: La iniciación La elongación La terminación. Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada. El ciclo de replicación de los virus es el término utilizado para describir el ciclo de reproducción de los virus. Este consta generalmente de las siguientes fases: fijación y entrada en la célula, eclipse, multiplicación y liberación del virus. Los virus son formas acelulares que no pueden reproducirse por si mismos en el exterior; para ello requieren introducirse en células donde controlarán sus mecanismos reproductivos.
Diapositiva2 ADHESION El primer paso en la infección de una célula es la adhesión o fijación a la superficie celular. Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula. Las células que carecen de los receptores apropiados no son susceptibles al virus.
Diapositiva3 PENETRACIÓN Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus. Virus envueltos (A) Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular. (B) Entada vía endosomas en la superficie celular. Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula. Virus no envueltos o desnudos Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
Diapositiva4 3. EMSAMBLAJE Y MADURACION Ensamblaje de los componentes de los virus para formar nuevas partículas víricas (proceso también llamado maduración). Los virus desnudos realizan su ensamblaje y maduración en forma intracelular y adquieren capacidad infectante dentro de la célula. Para su salida requieren de la desintegración celular. Los virus envueltos en el último paso al ensamblaje el virion se une con la membrana celular a la salida de la célula, mediante proteínas que se insertan en la superficie interna y exterior de la membrana. El virion naciente brota atraves de la membrana. El ensamblaje y maduración con la expulsión atraves de la superficie celular proporcionan un mecanismo más eficaz de salida, ya que no depende de la desintegración de la célula infectada.
Diapositiva5 SALIDA Liberación de los nuevos virus de la célula hospedadora, que ya pueden infectar otras células. Pueden producirse con o sin rotura de la célula, dependiendo de los virus. Los virus desnudos salen por lisis celular lo contrario de los envueltos que salen por gemación. La distribución en el cuerpo del hospedador de ciertos tipos de receptores celulares explica el tropismo tisular y de hospedador de los virus. Los tejidos y células que carecen de receptores específicos de virus determinados no son infectados por dichos viriones. La naturaleza de los receptores es variable y ciertos virus pueden tener más de un tipo de receptor.
1. ADSORPCIÓN El primer paso en la infección de una célula es la adhesión o fijación a la superficie celular. Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula. Las células que carecen de los receptores apropiados no son susceptibles al virus.
2. PENETRACIÓN Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus.
Virus envueltos
(A) Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular. (Figura 1).
(B) Entada vía endosomas en la superficie celular (figura 2)
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
Virus no envueltos o desnudos Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
3. PÉRDIDA DE LA CÁPSULA El ácido nucleido debe de estar lo suficientemente expuesto como para que la replicación viral comience en esta fase. Cuando el ácido nucleico es expuesto, las partículas víricas infecciosas no pueden removerse de las células – este es el comienz de la fase de ECLIPSE – la cual perdura hasta que viriones infecciosos nuevos sean creados.
4. SÍNTESIS DE ÁCIDO NUCLEICO Y PROTEINAS VIRALES Existen diversos mecanismos, algunos seran abarcados en capítulos posteriores.
5. ENSAMBLAJE/MADURACIÓN Nuevas partículas víricas son ensambladas. Puede haber un estadio de maduración posterior al proceso inicial de ensamblaje.
6. LIBERACIÓN O DESCARGA Los virus pueden ser liberados mediante lisis celular, o, si son envueltos, pueden yemar de las células. Los virus que yeman (figuras 3 y 4) no necesariamente destruyen la célula. Por tanto, algunos de estos virus son capaces de instaurar infecciones persistentes. No todas las partículas víricas liberadas son infecciosas. La proporción partículas infecciosas:partículas no-infecciosas varía según el virus y las condiciones de crecimiento.
El primer paso en la infección viral es la absorción a la membrana de la célula susceptible por medio de la adhesión de ligandos virales (proteínas de la capside o glucoproteínas de las espículas, por ejemplo a receptores superficiales de la célula). La distribución en el cuerpo del hospedador de ciertos tipos de receptores celulares explica el tropismo tisular y de hospedador de los virus.
Los tejidos y células que carecen de receptores específicos de virus determinados no son infectados por dichos viriones. La naturaleza de los receptores es variable y ciertos virus pueden tener más de un tipo de receptor. 2- Los virus complejos producen una rotura en la membrana celular del hospedador en uno de los puntos de anclaje, gracias a la presencia de algunas moléculas de enzimas hidrolíticas entre lasproteínas de la cápsida.
A través de la rotura, el tubo central inyecta el ADN vírico, quedando la cápside vacía en el exterior de la célula diana y el ácido nucleico libre en el citoplasma. La presencia de cápsidas en la superficie celular es un buen indicio de que ha sufrido una infección vírica.
Otros virus sin envoltura lipídica se introducen en la célula con cápsida y todo, lo cual puede realizarse de dos maneras: • Por penetración directa: después de la fijación, el virus abre una brecha en la membrana y se introduce en el citoplasma. • Por endocitosis: la membrana forma una invaginación en torno al virus, llegando a formar una vesícula que penetra en la célula.
Formada la vesícula, el virus abre una brecha en la membrana de la misma con ayuda de algunas enzimas hidrolíticas que él mismo transporta, penetrando así en el citoplasma
los virus con envoltura lipídica burlan la barrera de la membrana celular porque su cubierta lipídica se funde con la membrana, ya que son de la misma naturaleza. Esta fusión de membranas puede realizarse en dos lugares distintos: • Fusión en la superficie celular: de manera que el virión penetra directamente en el citoplasma. • Fusión con un lisosoma: se forma una vesícula por endocitosis, a la que se une un lisosoma para digerir la partícula introducida; entonces, la cubierta lipídica del virus se funde con la membrana del lisosoma y el virión escapa hacia el citoplasma.
3- Según la duración de la fase de eclipse, se suelen distinguir dos modalidades de ciclo infeccioso de un virus: • Ciclo ordinario: el ácido nucleico vírico procede inmediatamente a la transcripción de su mensaje genético en los ARN necesarios para su multiplicación, y prosigue rápidamente el ciclo vital. Este tipo de ciclo es el más extendido en la naturaleza. • Ciclo lisogénico: fue descubierto por André Lwoff en bacteriófagos. El ADN vírico se cierra por sus extremos generando un ADN circular. Este ADN se inserta en el ADN bacteriano en un lugar específico en el que la secuencia de nucleótidos bacterianos es semejante alguna región del ADN vírico.
Algunos virus que infectan células animales siguen también el ciclo lisogénico, como los papilomavirus de las verrugas y algunos retrovirus que producen algunos tipos de cáncer. En el caso de los retrovirus, conviene recordar que el ácido nucleico es ARN monocatenario, por lo que la transcriptasa inversa ha de copiar el genoma vírico en forma de ADN antes de que pueda insertarse en el ADN celular.
4- La multiplicación del virus consta de la replicación de su material genético, de la transcripción de su mensaje en una molécula de ARN y de la traducción del mensaje para producir proteínasvíricas, tanto las que formarán parte de la cápsida como las proteínas enzimáticas necesarias para el ensamblaje de las piezas del virión y para algunas de las funciones anteriores.
Losribosomas y la mayor parte de las enzimas que los ácidos nucleicos víricos utilizan en estos procesos son los de la célula infectada.
El primer paso en la infección de una célula es la adhesión o fijación a la superficie celular. Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura.
La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula.
Las células que carecen de los receptores apropiados no son susceptibles al virus.
Ensamblaje de los componentes de los virus para formar nuevas partículas víricas (proceso también llamado maduración).
Los virus desnudos realizan su ensamblaje y maduración en forma intracelular y adquieren capacidad infectante dentro de la célula. Para su salida requieren de la desintegración celular.
Los virus envueltos en el último paso al ensamblaje el virion se une con la membrana celular a la salida de la célula, mediante proteínas que se insertan en la superficie interna y exterior de la membrana.
El virion naciente brota atraves de la membrana. El ensamblaje y maduración con la expulsión atraves de la superficie celular proporcionan un mecanismo más eficaz de salida, ya que no depende de la desintegración de la célula infectada.
Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus.
Virus envueltos (A) Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática.
Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular.
(B) Entada vía endosomas en la superficie celular. Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular.
Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas.
A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma.
Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
Virus no envueltos o desnudos Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas.
Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
Diapositiva5 SALIDA Liberación de los nuevos virus de la célula hospedadora, que ya pueden infectar otras células.
Pueden producirse con o sin rotura de la célula, dependiendo de los virus.
Los virus desnudos salen por lisis celular lo contrario de los envueltos que salen por gemación. La distribución en el cuerpo del hospedador de ciertos tipos de receptores celulares explica el tropismo tisular y de hospedador de los virus.
Los tejidos y células que carecen de receptores específicos de virus determinados no son infectados por dichos viriones.
La naturaleza de los receptores es variable y ciertos virus pueden tener más de un tipo de receptor.
1 Etapas de la replicacion Viral Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada.
2 Adhesion El primer paso en la infección de una célula es la adhesión o fijación a la superficie celular. Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula. Las células que carecen de los receptores apropiados no son susceptibles al virus.
3 Penetracion Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus. Virus envueltos (A) Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular. (B) Entada vía endosomas en la superficie celular. Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula. Virus no envueltos o desnudos Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
4 Penetracion de los Bacteriografos Los virus complejos producen una rotura en la membrana celular del hospedador en uno de los puntos de anclaje, gracias a la presencia de algunas moléculas de enzimas hidrolíticas entre las proteínas de la cápside. A través de la rotura, el tubo central inyecta el ADN vírico, quedando la cápside vacía en el exterior de la célula diana y el ácido nucleico libre en el citoplasma. La presencia de cápsidas en la superficie celular es un buen indicio de que ha sufrido una infección vírica. Otros virus sin envoltura lipídica se introducen en la célula con cápsida y todo, lo cual puede realizarse de dos maneras: Por penetración directa: después de la fijación, el virus abre una brecha en la membrana y se introduce en el citoplasma. Por endocitosis: la membrana forma una invaginación en torno al virus, llegando a formar una vesícula que penetra en la célula. Formada la vesícula, el virus abre una brecha en la membrana de la misma con ayuda de algunas enzimas hidrolíticas que él mismo transporta, penetrando así en el citoplasma.
Los virus con envoltura lipídica burlan la barrera de la membrana celular porque su cubierta lipídica se funde con la membrana, ya que son de la misma naturaleza. Esta fusión de membranas puede realizarse en dos lugares distintos: Fusión en la superficie celular: de manera que el virión penetra directamente en el citoplasma.
Fusión con un lisosoma: se forma una vesícula por endocitosis, a la que se une un lisosoma para digerir la partícula introducida; entonces, la cubierta lipídica del virus se funde con la membrana del lisosoma y el virión escapa hacia el citoplasma.
5 Replicacion y Maduracion Ensamblaje de los componentes de los virus para formar nuevas partículas víricas (proceso también llamado maduración). Los virus desnudos realizan su ensamblaje y maduración en forma intracelular y adquieren capacidad infectante dentro de la célula. Para su salida requieren de la desintegración celular. Los virus envueltos en el último paso al ensamblaje el virion se une con la membrana celular a la salida de la célula, mediante proteínas que se insertan en la superficie interna y exterior de la membrana. El virion naciente brota atraves de la membrana. El ensamblaje y maduración con la expulsión atraves de la superficie celular proporcionan un mecanismo más eficaz de salida, ya que no depende de la desintegración de la célula infectada.
6 Maduracion y salida de virus desnudos y envueltos Después de la multiplicación del virus tiene lugar la salida de los nuevos individuos, que saldrán con capacidad de infectar nuevas células. Las principales modalidades de liberación da nombre a nuevas variantes del ciclo vital de los virus:
Infección persistente Los nuevos virus no esperan a la muerte de la célula hospedadora para abandonarla, sino que van saliendo de la célula al mismo tiempo que se van produciendo, de manera que la célula puede seguir viva y produciendo nuevas partículas víricas. La liberación puede hacerse de dos maneras: Los virus sin envoltura lipoproteica salen directamente, sin arrastrar ningún resto de la membrana plasmática, bien sea abriendo una brecha en la membrana, o bien aprovechando los mecanismos de exocitosis o salida de sustancias al exterior de la célula.
7 Salida de virus envuelto Liberación de los nuevos virus de la célula hospedadora, que ya pueden infectar otras células. Pueden producirse con o sin rotura de la célula, dependiendo de los virus. Los virus desnudos salen por lisis celular lo contrario de los envueltos que salen por gemación.
La replicación viral se divide en diversas fases o etapas y son: 1) reconocimiento y fijación a la célula hospedadora, mediado por receptores específicos; 2) penetración del virus al interior de la célula mediante endocitosis o fusión de membranas; 3) pérdida de la cápside y liberación del genoma viral; 4) síntesis precoz de RNAm viral y de proteínas virales no estructurales necesarias para la replicación y la transcripción viral; 5) replicación del genoma viral; 6) síntesis tardía de RNAm viral y de proteínas estructurales; 7) modificación postranscripcional de las proteínas virales; 8) ensamblaje y formación de las partículas virales, que se puede realizar en distintas partes de la célula (núcleo, citoplasma, membrana plasmática, etc.); 9) liberación del virus mediante lisis celular, exocitosis o gemación; 10) maduración de la partícula viral (partícula infectiva) 1.- Fijación, penetración y pérdida de la cubierta (iniciación) • El virus se fija o adsorbe a un sitio de la superficie celular que actúan como receptores específicos. • Después entra a la célula (penetración) • Y por último pierde la cubierta poco después de haber ingresado a la célula 2.- Síntesis de los componentes vitales (replicación) • Inicia la fase de sinética de la replicación viral • Se sintetizan todas las macromoléculas especificas del virus. • Las proteínas son sintetizadas en el citoplasma • El DNA suele replicarse en el núcleo, mientras que el ARN viral se duplica en el citoplasma (existen excepciones) 3.- Morfogénesis y liberación (fase de liberación) • Formación de los virus descendientes • Adquisición de la cubierta • Muerte de la célula hospedadora (varia) Los virus cuentan con 2 ciclos de replicación. Algunos virus se pueden replicar por lo que se conoce como ciclo Lítico; ellos inyectan a la célula huésped con su ADN, obligándola a fabricar nuevos virus, hasta que la célula huésped explota liberando los patógenos al medio.Otros virus operan de diferente forma: ellos entran e inyectan su ADN en la célula huésped pero, en lugar de tomar el control y fabricar mas virus, el ADN inyectado puede tornarse inactivo por un cierto tiempo, hasta que un apropiado evento celular dispara el proceso nuevamente. A éste ciclo se le conoce como Lisogènico.El bacteriófago, también conocido como fago, es un elemento extra cromosomal del estudio de las bacterias, en otras palabras, es un virus capaz de infectar a células eucariontes y procariontes, son capaces de replicarse por si solos, portan información genética y pueden encontrarse en casi todos los lugares pero por lo general se localizan donde están sus huéspedes obligatorios. Se reproduce en células procariontes.Esta formado por: Cabeza, cuello, cola, Vaina, placa basal, fibras de la cola y espículas.El bacteriófago también lleva a cabo un ciclo de replicación el cual se puede dividir en distintas etapas, las cuales son muy comunes a otros virus bacterianos y eucarióticos. Replicación de los virus ADN La mayoría de los virus ADN codifican su propia polimerasa ADN, excepto los virus ADN pequeños, como los parvovirus, papillomavirus. Esta diferencia, significa que los virus ADN pequeños necesitan células huésped en fase mitótica para suministrarles la ADN polimerasa celular Hay 3 maneras en que los virus ADN replican su material genético: Replicación bidireccional a partir de un substrato circular. En este caso, hay una síntesis simultánea de las cadenas ADN líder y de cierre, empezando en un origen definido de la replicación de ADN. Ejem. Los papovavirus. Replicación desde un substrato linear. En este caso, la síntesis de las nuevas cadenas de ADN no es simultánea. Ocurre secuencialmente (primero se hace una cadena en su totalidad, luego la siguiente, etc). Un ejemplo: los adenovirus. Replicación por medio de un intermediario ARN. Un ejemplo de este sistema son los hepaADNvirus (HBV), los cuales tienen parcialmente un genoma dsADN que debe ser convertido en un complejo ARN por la enzima transcriptasa inversa viral, durante el ciclo vital del virus.
1. ADSORPCIÓN El primer paso en la infección de una célula es la adhesión o fijación a la superficie celular. Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula. Las células que carecen de los receptores apropiados no son susceptibles al virus.
2. PENETRACIÓN Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus.
Virus envueltos
(A) Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular. (Figura 1).
(B) Entada vía endosomas en la superficie celular (figura 2)
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
Virus no envueltos o desnudos Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
3. PÉRDIDA DE LA CÁPSULA El ácido nucleido debe de estar lo suficientemente expuesto como para que la replicación viral comience en esta fase. Cuando el ácido nucleico es expuesto, las partículas víricas infecciosas no pueden removerse de las células – este es el comienz de la fase de ECLIPSE – la cual perdura hasta que viriones infecciosos nuevos sean creados.
4. SÍNTESIS DE ÁCIDO NUCLEICO Y PROTEINAS VIRALES Existen diversos mecanismos, algunos seran abarcados en capítulos posteriores.
5. ENSAMBLAJE/MADURACIÓN Nuevas partículas víricas son ensambladas. Puede haber un estadio de maduración posterior al proceso inicial de ensamblaje.
6. LIBERACIÓN O DESCARGA Los virus pueden ser liberados mediante lisis celular, o, si son envueltos, pueden yemar de las células. Los virus que yeman (figuras 3 y 4) no necesariamente destruyen la célula. Por tanto, algunos de estos virus son capaces de instaurar infecciones persistentes. No todas las partículas víricas liberadas son infecciosas. La proporción partículas infecciosas:partículas no-infecciosas varía según el virus y las condiciones de crecimiento.
espués de la multiplicación del virus tiene lugar la salida de los nuevos individuos, que saldrán con capacidad de infectar nuevas células. Las principales modalidades de liberación da nombre a nuevas variantes del ciclo vital de los virus: Infección persistente[editar · editar código] Los nuevos virus no esperan a la muerte de la célula hospedadora para abandonarla, sino que van saliendo de la célula al mismo tiempo que se van produciendo, de manera que la célula puede seguir viva y produciendo nuevas partículas víricas. La liberación puede hacerse de dos maneras: Los virus sin envoltura lipoproteica salen directamente, sin arrastrar ningún resto de la membrana plasmática, bien sea abriendo una brecha en la membrana, o bien aprovechando los mecanismos de exocitosis o salida de sustancias al exterior de la célula. Los virus con envoltura lipoproteica salen por gemación, es decir, se rodean de una porción de membrana plasmática que acaba separándose de la célula y constituye la cubierta lipoproteica del nuevo virus.
1- El primer paso en la infección viral es la absorción a la membrana de la célula susceptible por medio de la adhesión de ligandos virales (proteínas de la capside o glucoproteínas de las espículas, por ejemplo a receptores superficiales de la célula). La distribución en el cuerpo del hospedador de ciertos tipos de receptores celulares explica el tropismo tisular y de hospedador de los virus. Los tejidos y células que carecen de receptores específicos de virus determinados no son infectados por dichos viriones. La naturaleza de los receptores es variable y ciertos virus pueden tener más de un tipo de receptor.
2- Los virus complejos producen una rotura en la membrana celular del hospedador en uno de los puntos de anclaje, gracias a la presencia de algunas moléculas de enzimas hidrolíticas entre lasproteínas de la cápsida. A través de la rotura, el tubo central inyecta el ADN vírico, quedando la cápside vacía en el exterior de la célula diana y el ácido nucleico libre en el citoplasma. La presencia de cápsidas en la superficie celular es un buen indicio de que ha sufrido una infección vírica.
Otros virus sin envoltura lipídica se introducen en la célula con cápsida y todo, lo cual puede realizarse de dos maneras:
• Por penetración directa: después de la fijación, el virus abre una brecha en la membrana y se introduce en el citoplasma. • Por endocitosis: la membrana forma una invaginación en torno al virus, llegando a formar una vesícula que penetra en la célula. Formada la vesícula, el virus abre una brecha en la membrana de la misma con ayuda de algunas enzimas hidrolíticas que él mismo transporta, penetrando así en el citoplasma. Los virus con envoltura lipídica burlan la barrera de la membrana celular porque su cubierta lipídica se funde con la membrana, ya que son de la misma naturaleza. Esta fusión de membranas puede realizarse en dos lugares distintos: • Fusión en la superficie celular: de manera que el virión penetra directamente en el citoplasma. • Fusión con un lisosoma: se forma una vesícula por endocitosis, a la que se une un lisosoma para digerir la partícula introducida; entonces, la cubierta lipídica del virus se funde con la membrana del lisosoma y el virión escapa hacia el citoplasma.
3- Según la duración de la fase de eclipse, se suelen distinguir dos modalidades de ciclo infeccioso de un virus: • Ciclo ordinario: el ácido nucleico vírico procede inmediatamente a la transcripción de su mensaje genético en los ARN necesarios para su multiplicación, y prosigue rápidamente el ciclo vital. Este tipo de ciclo es el más extendido en la naturaleza. • Ciclo lisogénico: fue descubierto por André Lwoff en bacteriófagos. El ADN vírico se cierra por sus extremos generando un ADN circular. Este ADN se inserta en el ADN bacteriano en un lugar específico en el que la secuencia de nucleótidos bacterianos es semejante alguna región del ADN vírico.
Algunos virus que infectan células animales siguen también el ciclo lisogénico, como los papilomavirus de las verrugas y algunos retrovirus que producen algunos tipos de cáncer. En el caso de los retrovirus, conviene recordar que el ácido nucleico es ARN monocatenario, por lo que la transcriptasa inversa ha de copiar el genoma vírico en forma de ADN antes de que pueda insertarse en el ADN celular.
4- La multiplicación del virus consta de la replicación de su material genético, de la transcripción de su mensaje en una molécula de ARN y de la traducción del mensaje para producir proteínasvíricas, tanto las que formarán parte de la cápsida como las proteínas enzimáticas necesarias para el ensamblaje de las piezas del virión y para algunas de las funciones anteriores. Losribosomas y la mayor parte de las enzimas que los ácidos nucleicos víricos utilizan en estos procesos son los de la célula infectada.
El primer paso en la infección de una célula es la adhesión o fijación a la superficie celular. Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula.
Las células que carecen de los receptores apropiados no son susceptibles al virus.
ETAPAS DE LA REPLICACION VIRAL Slide 1: Etapas de la replicación viral 2012-0593 El ciclo de replicación de los virus es el término utilizado para describir el ciclo de reproducción de los virus. Este consta generalmente de las siguientes fases: fijación y entrada en la célula, eclipse, multiplicación y liberación del virus. Los virus son formas acelulares que no pueden reproducirse por si mismos en el exterior; para ello requieren introducirse en células donde controlarán sus mecanismos reproductivos Slide 2: Absorcion 2012-0593 El primer paso en la infección de una célula es la adhesión o fijación a la superficie celular. Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula. Las células que carecen de los receptores apropiados no son susceptibles al virus. Slide 3: Penetracion 2012-0593 La penetración a la célula blanco ocurre normalmente en un tiempo muy breve luego de la unión del virus al receptor en la superficie celular. A diferencia del proceso anterior, el ingreso a la célula es generalmente un proceso dependiente de energía, esto significa que la célula debe estar metabolicamente activa . En este paso hay tres mecanismos involucrados: 1) TRANSLOCACIÓN: La partícula viral atraviesa la membrana citoplasmática. Este proceso es poco frecuente en los virus y no está completamente entendido. Puede estar mediado por proteínas de la cápside viral y receptores específicos de membrana 2) ENDOCITOSIS: El virus ingresa en vacuolas intracelulares. Este es probablemente el mecanismo de ingreso más común en los virus. No se necesita de la presencia de proteínas virales específicas -excepto de aquellas involucradas en la unión al receptor- pero si es necesaria la formación de vesículas endocíticas en la membrana celular y la posterior internalización de estas. Este mecanismo involucra un cambio de pH en el endosoma para liberar posteriormente el genoma del virus dentro del citoplasma. 3) FUSIÓN: Sólo ocurre con los virus envueltos. En este proceso ocurre una fusión de la envoltura viral con la membrana celular, ya sea directamente en la superficie celular o en una vesícula citoplasmática luego de la endocitosis. La fusión requiere de la presencia de una proteína de fusión específica en la envoltura del virus -por ejemplo la hemaglutinina en el virus de la influenza o las glucoproteínas de transmembrana en los retrovirus-. Estas proteínas promueven la unión de las membranas del virus y de la célula, esto resulta en la liberación de la nucleocápside directamente en el citoplasma. Este proceso no es dependiente pH. El proceso de endocitosis es casi universal en las células animales; se forman vesículas en la membrana plasmática, las que posteriormente son englobadas e internalizadas al citoplasma. La vida media de estas vesículas es muy breve, en pocos segundos la mayoría se fusionan con endosomas liberando su contenido en esas vesículas más grandes. Hasta este momento cualquier virus contenido en estas estructuras, aún está separado del citoplasma por una bicapa lipídica , por lo tanto no se puede decir estrictamente que ha ingresado en la célula. Posteriormente, los endosomas se fusionan con los lisosomas, el ambiente interno se vuelve progresivamente más hostil, se acidifica en la medida que el pH desciende y aumenta la concentración de enzimas degradativas . Esto significa que el virus debe liberarse de la vesícula e ingresar al citoplasma antes de ser degradado. Hay una serie de mecanismos por los cuales esto ocurre, incluyendo la fusión de membranas. La liberación de la partícula viral desde el endosoma y su pasaje al citoplasma están conectados estrechamente -y a menudo imposible de separar- del proceso de liberación de las estructuras, liberación del genoma -.
Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus.
Virus envueltos
(A) Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular.
(B) Entada vía endosomas en la superficie celular.
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
Virus no envueltos o desnudos Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
Slide 4: Penetracion Bacteriofago 2012-0593 La primera etapa de la infección viral es la interacción de un virión con un sitio receptor específica de la superficie celular. Las moléculas receptoras difieren para los distintos virus, y son proteínas en algunos casos (por ejemplo, picornavirus) y oligosacáridos en otros (por ejemplo, ortovirus y paramixovirus). La presencia o la ausencia de receptores desempeña una función determinante de primer orden en el tropismo celular y la patogenia viral; por ejemplo, el poliovirus es capaz de fijarse sólo en las células del sistema nervioso central y las vías digestivas de los primates. Se cree que la fijación en receptores refleja homologías configurativas fortuitas entre la estructura de la superficie del virión y un componente de la superficie celular. Por ejemplo, el virus de la inmunodeficiencia humana (HIV) se une al receptor CD4 en las células del sistema inmunitario y se cree que el virus de la rabia interactúa con los receptores de acetilcolina y que el virus de Epstein-Barr reconoce al receptor para el tercer componente del complemento en las células B. Es probable que cada célula susceptible posea por lo menos 100,000 sitios receptores para un virus dado Slide 5: Replicacion y Maduracion 2012-0593 Este es el estadío del ciclo replicativo en el cual el virus se hace infeccioso. La maduración generalmente incluye cambios estructurales en la partícula viral, estos cambios pueden ser el resultado de cortes específicos de proteínas de la cápside para formar productos maduros, o de cambios conformacionales en las proteínas durante el ensamble. Estos eventos conducen frecuentemente a cambios estructurales sustanciales en la cápside, por ej. las diferencias de antigenicidad entre la partícula viral incompleta y la madura. Las proteasas virales frecuentemente están involucradas en la maduración, aunque en algunos casos pueden actuar enzimas celulares o una mezcla de enzimas celulares y virales. Ciertamente hay un cierto riesgo en la participación de enzimas proteolíticas celulares en este proceso, dado que la falta de especificidad sobre el sustrato podría ocasionar fácilmente la completa degradación de las proteínas de la cápside. Sin embargo las proteasas codificadas por el virus, en general son altamente específicas para determinadas secuencias de aminoácidos y estructuras; frecuentemente sólo cortan una unión peptídica en particular en una estructura proteica grande y compleja como la cápside. Además, en cierta forma son controladas al ser empaquetadas dentro de la partícula viral durante el ensamble y sólo se activan cuando toman contacto con las secuencias específicas para la conformación de la cápside, por ejemplo al localizarse en un sitio hidrofóbico o por cambios de pH o concentración de determinados iones dentro de la cápside. Las proteasas de los retrovirus son un buen ejemplo de enzimas involucradas en la maduración, que están bajo este tipo de control. El core de los retrovirus está compuesto por proteínas codificadas por el gen gag y la proteasa es empaquetada en el core antes de la liberación de la célula por gemación. En algún momento del proceso de gemación, la proteasa corta los precursores de la proteína gag en productos maduros -proteínas de la càpside, nucleocàpside y matriz de la partícula viral madura-. Para algunos virus el ensamble y la maduración ocurren dentro de la célula y son procesos inseparables, mientras que para otros la maduración puede ocurrir sólo después de la liberación de la partícula viral. De todos modos, y en cualquiera de los casos, el proceso de maduración prepara al virus para la infección de nuevas células. En resumen, la replicación de los virus implica tres procesos básicos: a) El inicio de la infección. b) La replicación y expresión del genoma. c) La liberación del virión maduro desde la célula infectada
Slide 6: Maduracion y Lberacion 2012-0593 Los virus en general pueden liberarse de la célula mediante dos tipos de mecanismos: lisis y gemación. Para los virus líticos -la mayoría de los virus desnudos-, la liberación es un proceso simple: la célula infectada se rompe y libera las partículas virales. En el caso de los virus que se liberan por gemación, estos adquieren su envoltura lipídica y las proteínas asociadas en el momento en que pasan a través de la membrana plasmática o de vesículas intracelulares. La liberación de los virus por esta vía puede resultar altamente perjudicial para la célula -por ej.en el caso de paramixovirus, rabdovirus, togavirus- o no -por ej. retrovirus-; de todos modos en cualquiera de los casos el proceso es controlado por el virus: la interacción física de las proteínas de la cápside sobre la superficie interna de la membrana celular fuerza a la partícula viral a pasar a través de dicha membrana. El ensamble, la maduración y liberación son generalmente procesos simultáneos en los virus que se liberan por gemación. El tipo de membrana a través de la cual geman los virus depende del tipo de virus, por ej. puede ser a través de la membrana celular como en el caso de los retrovirus y togavirus, a través de la membrana citoplasmática -ortomixovirus, coronavirus, bunyavirus, rabdovirus, hepadnavirus- o a través de la membrana nuclear como en los herpesvirus. Slide 7: Salida de un virus envuelto 2012-0593 Se encuentra la variación más amplia en las estrategias de la expresión genética entre los virus que contienen ARN. Algunos viriones poseen polimerasas (ortomixovirus, reovirus); otros sistemas emplean mensajes subgenómicos generados a veces por empalme (ortomixovirus, retrovirus); por último, algunos virus sintetizan grandes precursores poliproteínicos que se someten a procesamiento y segmentación para generar los productos génicos finales (picornavirus, retrovirus).Los sitios intracelulares en los que ocurren los diferentes acontecimientos de la replicación viral, varían de un grupo a otro. Es posible plantear algunas generalizaciones. La proteína viral se sintetiza en el citoplasma, sobre los polirribosomas compuestos por ARNm específico del virus y ribosomas de la célula huésped. El ADN viral suele replicarse en el núcleo. El ARN genómico viral se duplica generalmente en el citoplasma celular, aunque hay excepciones
La primera etapa de la infección viral es la interacción de un virión con un sitio receptor específica de la superficie celular.
Las moléculas receptoras difieren para los distintos virus, y son proteínas en algunos casos (por ejemplo, picornavirus) y oligosacáridos en otros (por ejemplo, ortovirus y paramixovirus).
La presencia o la ausencia de receptores desempeña una función determinante de primer orden en el tropismo celular y la patogenia viral; por ejemplo, el poliovirus es capaz de fijarse sólo en las células del sistema nervioso central y las vías digestivas de los primates. Se cree que la fijación en receptores refleja homologías configurativas fortuitas entre la estructura de la superficie del virión y un componente de la superficie celular. Por ejemplo, el virus de la inmunodeficiencia humana (HIV) se une al receptor CD4 en las células del sistema inmunitario y se cree que el virus de la rabia interactúa con los receptores de acetilcolina y que el virus de Epstein-Barr reconoce al receptor para el tercer componente del complemento en las células B.
Es probable que cada célula susceptible posea por lo menos 100,000 sitios receptores para un virus dado.
mediante Después de la fijación, la partícula viral entra en la célula. Esta etapa se conoce como penetración, viropexia o fagocitosis.
En algunos sistemas, se realiza por endocitosis mediada por receptores, con captación de las partículas virales ingeridas dentro de endosomas.
Están menos claros los detalles de la penetración en otros sistemas. Ocurre perdida de la cubierta de manera concomitante con la penetración o poco después de la misma. La pérdida de la cubierta, o descubrimiento, es la separación física del ácido nucleico viral (o, en algunos casos, de las nucleocápsides inteARNs) a partir de los componentes estructurales externos del virión. En este momento se pierde la infecciosidad del virus original.
Los virus son los únicos agentes infecciosos en los que la disolución del agente infectante es una etapa obligatoria de la vía de la replicación
Maduracion y salida de virus desnudos y envueltos Después de la multiplicación del virus tiene lugar la salida de los nuevos individuos, que saldrán con capacidad de infectar nuevas células. Las principales modalidades de liberación da nombre a nuevas variantes del ciclo vital de los virus:
Infección persistente Los nuevos virus no esperan a la muerte de la célula hospedadora para abandonarla, sino que van saliendo de la célula al mismo tiempo que se van produciendo, de manera que la célula puede seguir viva y produciendo nuevas partículas víricas
La liberación puede hacerse de dos maneras: Los virus sin envoltura lipoproteica salen directamente, sin arrastrar ningún resto de la membrana plasmática, bien sea abriendo una brecha en la membrana, o bien aprovechando los mecanismos de exocitosis o salida de sustancias al exterior de la célula.
Liberación de los nuevos virus de la célula hospedadora, que ya pueden infectar otras células. Pueden producirse con o sin rotura de la célula, dependiendo de los virus. Los virus desnudos salen por lisis celular lo contrario de los envueltos que salen por gemación. -todo Se encuentra la variación más amplia en las estrategias de la expresión genética entre los virus que contienen ARN. Algunos viriones poseen polimerasas (ortomixovirus, reovirus); otros sistemas emplean mensajes subgenómicos generados a veces por empalme (ortomixovirus, retrovirus); por último, algunos virus sintetizan grandes precursores poliproteínicos que se someten a procesamiento y segmentación para generar los productos génicos finales (picornavirus, retrovirus).
La extensión a la que las enzimas específicas del virus participan en estos procesos varía de un grupo a otro. En general, los virus de mayor tamaño (herpesvirus, poxvirus) son más independientes de las funciones celulares que los virus de menor tamaño. Esta es una razón de que los virus de mayor tamaño sean más sensibles a la quimioterapia antiviral, porque se dispone de más procesos específicos de los virus como blancos de la acción farmacológica.
Los sitios intracelulares en los que ocurren los diferentes acontecimientos de la replicación viral, varían de un grupo a otro. Es posible plantear algunas generalizaciones. La proteína viral se sintetiza en el citoplasma, sobre los polirribosomas compuestos por ARNm específico del virus y ribosomas de la célula huésped. El ADN viral suele replicarse en el núcleo. El ARN genómico viral se duplica generalmente en el citoplasma celular, aunque hay excepciones
Diapositiba1 Etapas de la replicación viral La replicación es el proceso en el cual se copia el ADN. Este proceso es semiconservativo y bidireccional. Funciona igual, tanto en procariontes como en eucarionte, salvo algunos cambio, principalmente de las proteínas que participan. Se dieron muchas hipótesis sobre como se dupllicaba el ADN hasta que Watson y Crick propusieron la hipótesis semiconservativa, según la cual, las nuevas moléculas de ADN formadas a partir de otra antigua, tienen una hebra antigua y otra nueva. La replicación ocurre en 3 etapas: La iniciación La elongación La terminación. Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada. El ciclo de replicación de los virus es el término utilizado para describir el ciclo de reproducción de los virus. Este consta generalmente de las siguientes fases: fijación y entrada en la célula, eclipse, multiplicación y liberación del virus. Los virus son formas acelulares que no pueden reproducirse por si mismos en el exterior; para ello requieren introducirse en células donde controlarán sus mecanismos reproductivos.
Diapositiva 2 ADHESION El primer paso en la infección de una célula es la adhesión o fijación a la superficie celular. Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula. Las células que carecen de los receptores apropiados no son susceptibles al virus.
2011-0029 José Luis González Salas Diapositiva 3 PENETRACIÓN Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus. Virus envueltos (A) Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular. (B) Entada vía endosomas en la superficie celular. Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula. Virus no envueltos o desnudos Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
Diapositiva 4 EMSAMBLAJE Y MADURACION Ensamblaje de los componentes de los virus para formar nuevas partículas víricas (proceso también llamado maduración). Los virus desnudos realizan su ensamblaje y maduración en forma intracelular y adquieren capacidad infectante dentro de la célula. Para su salida requieren de la desintegración celular. Los virus envueltos en el último paso al ensamblaje el virion se une con la membrana celular a la salida de la célula, mediante proteínas que se insertan en la superficie interna y exterior de la membrana. El virion naciente brota atraves de la membrana. El ensamblaje y maduración con la expulsión atraves de la superficie celular proporcionan un mecanismo más eficaz de salida, ya que no depende de la desintegración de la célula infectada.
2011-0029 José Luis González Salas Diapositiva5 SALIDA Liberación de los nuevos virus de la célula hospedadora, que ya pueden infectar otras células. Pueden producirse con o sin rotura de la célula, dependiendo de los virus. Los virus desnudos salen por lisis celular lo contrario de los envueltos que salen por gemación. La distribución en el cuerpo del hospedador de ciertos tipos de receptores celulares explica el tropismo tisular y de hospedador de los virus. Los tejidos y células que carecen de receptores específicos de virus determinados no son infectados por dichos viriones. La naturaleza de los receptores es variable y ciertos virus pueden tener más de un tipo de receptor.
2011-0029 José Luis González Salas Diapositiva 6 Maduracion y salida de virus desnudos y envueltos Después de la multiplicación del virus tiene lugar la salida de los nuevos individuos, que saldrán con capacidad de infectar nuevas células. Las principales modalidades de liberación da nombre a nuevas variantes del ciclo vital de los virus:
Infección persistente Los nuevos virus no esperan a la muerte de la célula hospedadora para abandonarla, sino que van saliendo de la célula al mismo tiempo que se van produciendo, de manera que la célula puede seguir viva y produciendo nuevas partículas víricas
La liberación puede hacerse de dos maneras: Los virus sin envoltura lipoproteica salen directamente, sin arrastrar ningún resto de la membrana plasmática, bien sea abriendo una brecha en la membrana, o bien aprovechando los mecanismos de exocitosis o salida de sustancias al exterior de la célula.
2011-0029 José Luis González Salas Diapositiva 7 SALIDA
Liberación de los nuevos virus de la célula hospedadora, que ya pueden infectar otras células. Pueden producirse con o sin rotura de la célula, dependiendo de los virus. Los virus desnudos salen por lisis celular lo contrario de los envueltos que salen por gemación. -todo Se encuentra la variación más amplia en las estrategias de la expresión genética entre los virus que contienen ARN. Algunos viriones poseen polimerasas (ortomixovirus, reovirus); otros sistemas emplean mensajes subgenómicos generados a veces por empalme (ortomixovirus, retrovirus); por último, algunos virus sintetizan grandes precursores poliproteínicos que se someten a procesamiento y segmentación para generar los productos génicos finales (picornavirus, retrovirus).
La extensión a la que las enzimas específicas del virus participan en estos procesos varía de un grupo a otro. En general, los virus de mayor tamaño (herpesvirus, poxvirus) son más independientes de las funciones celulares que los virus de menor tamaño. Esta es una razón de que los virus de mayor tamaño sean más sensibles a la quimioterapia antiviral, porque se dispone de más procesos específicos de los virus como blancos de la acción farmacológica.
Los sitios intracelulares en los que ocurren los diferentes acontecimientos de la replicación viral, varían de un grupo a otro. Es posible plantear algunas generalizaciones. La proteína viral se sintetiza en el citoplasma, sobre los polirribosomas compuestos por ARNm específico del virus y ribosomas de la célula huésped. El ADN viral suele replicarse en el núcleo. El ARN genómico viral se duplica generalmente en el citoplasma celular, aunque hay excepciones
89693 - Fabiano Lima das Gracas Virologia – Grupo Lunes 10-12pm
1-Etapas de la Replicacion Viral: Ciclo de los virus • La adhesión o adsorción es una unión específica entre proteínas de la cápside vírica y receptores específicos de la superficie celular del huésped, pero algunos bacteriófagos también son capaces de adherirse a los flagelos, vellosidades (pili) o cápsulas presentes en la superficie de la bacteria hospedera. • La penetración sigue a la adhesión; los virus se introducen en la célula huésped mediante endocitosis mediada por receptores o por fusión de membrana. Esto recibe a menudo el nombre de penetración vírica • El despojo es el proceso en que la cápside vírica es degradada por enzimas virales o del huésped, liberando así el ácido nucleico del genoma vírico. • La replicación implica la síntesis de ARN mensajero (ARNm) vírico en todos los virus con rasgos de ARN positivos, la síntesis de proteínas víricas, el ensamblaje de proteínas víricas y la replicación del genoma viral • Tras el ensamblaje de partículas víricas, a menudo se produce una modificación postraduccional de las proteínas víricas.
89693 - Fabiano Lima das Gracas Virologia – Grupo Lunes 10-12pm
2-Adhesion:
El primer paso en la infección de una célula es la adhesión o fijación a la superficie celular. Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula. Las células que carecen de los receptores apropiados no son susceptibles al virus.
89693 - Fabiano Lima das Gracas Virologia – Grupo Lunes 10-12pm
3-Penetracion:
Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus. Virus envueltos • Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular. • Entada vía endosomas en la superficie celular Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula. Virus no envueltos o desnudos Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
89693 - Fabiano Lima das Gracas Virologia – Grupo Lunes 10-12pm
4-Penetracion de los Bacteriofagos:
Los bacteriófagos (también llamados fagos), son virus que infectan exclusivamente a bacterias. Para entrar en una célula, los fagos se acoplan a receptores específicos en la superficie de la bacteria, que pueden ser lipopolisacáridos, ácidos teicoicos, proteínas o incluso flagelos. Por ello, cada fago solo podrá infectar ciertas bacterias según sus receptores. Puesto que los fagos no son móviles, dependen de encuentros al azar con los receptores adecuados en solución para poder infectar una bacteria. Parece que los bacteriófagos presentan una especie de jeringa mediante la cual introducen su material genético en el interior de la célula. Tras el reconocimiento del receptor adecuado, la cola y cuello del fago se contraen, quedando así el fago acoplado a la superficie celular. El material genético puede ser ahora introducido a través de la membrana o bien simplemente depositado sobre la superficie. No se descarta que pueda haber fagos con otros métodos diferentes para introducir su material genético en la célula.
89693 - Fabiano Lima das Gracas Virologia – Grupo Lunes 10-12pm
5-Replicacion y Maduracion:
Los genomas virales recién sintetizados y los polipéptidos de la cápside se ensamblan para formar los virus descendientes. Las cápsides icosaédricas pueden condensarse en ausencia de ácido nucleico, en tanto que las nucleocápsides de los virus de simetría helicoidal no pueden formarse sin ARN viral. No existen mecanismos especiales para la liberación de virus carentes de cubierta; las células infectadas acaban por experimentar lisis y liberar las partículas virales. Los virus cubiertos maduran por un proceso de gemación. Se insertan glucoproteínas de cubierta específica del virus en las membranas celulares y, a continuación, las nucleocápsides hacen gemación a través de la membrana a nivel de estos sitios modificados y, al hacerlo así, adquieren su cubierta. La gemación ocurre a menudo a nivel de la membrana plasmática y puede abarcar a otras membranas de la célula. Los virus cubiertos no son infecciosos hasta que han adquirido sus cubiertas. Por lo tanto, no se acumulan de manera típica viriones descendientes infecciosos en el interior de la célula infectada. La maduración viral es a veces un proceso ineficaz. Pueden acumularse cantidades excesivas de componentes virales y participar en la formación de cuerpos de inclusión en la célula Como resultado de los profundos efectos dañinos de la replicación viral, se desarrollan por último efectos citopáticos celulares y la célula muere. Sin embargo, hay casos en que el virus no lesiona a la célula y se producen infecciones persistentes y prolongadas.
89693 - Fabiano Lima das Gracas Virologia – Grupo Lunes 10-12pm
6-Maduracion y Salida de Virus Desnudos y Envueltos:
Virus envueltos • Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular. • Entada vía endosomas en la superficie celular Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula. Virus no envueltos o desnudos Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
89693 - Fabiano Lima das Gracas Virologia – Grupo Lunes 10-12pm
7-Salida de un Virus Envuelto:
Los virus pueden ser liberados mediante lisis celular, o, si son envueltos, pueden yemar de las células. Los virus que yeman no necesariamente destruyen la célula. Por tanto, algunos de estos virus son capaces de instaurar infecciones persistentes. No todas las partículas víricas liberadas son infecciosas. La proporción partículas infecciosas:partículas no-infecciosas varía según el virus y las condiciones de crecimiento.
88977 ETAPAS DE REPLICACION VIRAL El ciclo de replicación de los virus es el término utilizado para describir el ciclo de reproducción de los virus. Este consta generalmente de las siguientes fases: fijación y entrada en la célula, eclipse, multiplicación y liberación del virus. Los virus son formas acelulares que no pueden reproducirse por si mismos en el exterior; para ello requieren introducirse en células donde controlarán sus mecanismos
88977 ADHESION El primer paso en la infección de una célula es la adhesión o fijación a la superficie celular. Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula. Las células que carecen de los receptores apropiados no son susceptibles al virus.
88977 PENETRACIÓN Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus.
Virus envueltos
(A) Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular.
(B) Entada vía endosomas en la superficie celular.
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
Virus no envueltos o desnudos Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
88977 PENETRACION DE UN BACTERIOFAGO ciclo litico de un bacteriófago.formado por acido nuceico ADN y capsida compleja. fijacion o adsorcion: se fijan inicialmente a traves de las puntas de las fibras cudales mediante enlaces quimicos y posterormente de forma mecanica al clavar las pespinas basales en la pared bacteriana.
penetracion: el bacteriofago mediante enzimas lisozimas situadas en su placa basal perrfora la pared cecular y luego contrae la vaina e introduce el eje tubular de modo que el adn pasa directamente al citoplasma
88977 REPLICACION Y MADURACION Ensamblaje de los componentes de los virus para formar nuevas partículas víricas (proceso también llamado maduración). Los virus desnudos realizan su ensamblaje y maduración en forma intracelular y adquieren capacidad infectante dentro de la célula. Para su salida requieren de la desintegración celular. Los virus envueltos en el último paso al ensamblaje el virion se une con la membrana celular a la salida de la célula, mediante proteínas que se insertan en la superficie interna y exterior de la membrana. El virion naciente brota atraves de la membrana. El ensamblaje y maduración con la expulsión atraves de la superficie celular proporcionan un mecanismo más eficaz de salida, ya que no depende de la desintegración de la célula infectada
Salida de virus envuelto Liberación de los nuevos virus de la célula hospedadora, que ya pueden infectar otras células. Pueden producirse con o sin rotura de la célula, dependiendo de los virus. Los virus desnudos salen por lisis celular lo contrario de los envueltos que salen por gemación.Después de la multiplicación del virus tiene lugar la salida de los nuevos individuos, que saldrán con capacidad de infectar nuevas células. Las principales modalidades de liberación da nombre a nuevas variantes del ciclo vital de los virus:
Christopher Robles 2011-0692 Etapas de la replicacion viral
Slide 1 Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada.
La única característica de la multiplicación viral consiste en que, poco después de su interacción con una célula huésped, el virión infectante se desintegra y pierde la infecciosidad que se puede medir. Esta fase del ciclo celular se llama periodo de eclipse; su duración varía dependiendo del virus y de la célula huésped en particular, y termina con la formación de las primeras partículas virales descendientes infecciosas. El periodo de eclipse es, en realidad, de intensa actividad sintética, puesto que la célula se redirige hacia la satisfacción de las necesidades del "pirata" viral. En algunos casos, tan pronto como el ácido nucleico viral penetra a la célula huésped, el metabolismo celular es recanalizado exclusivamente hacia la síntesis de nuevas partículas virales. En otros casos, los procesos metabólicos de la célula huésped no se alteran significativamente, aunque la célula sintetiza proteínas virales y ácidos nucleicos.
Slide 2, Los virus han desarrollado diversas estrategias para lograr la multiplicación en las células huéspedes parasitadas por ellos. Aunque los detalles varían de un grupo a otro, el patrón general de los ciclos de replicación es semejante. La primera etapa de la infección viral es la interacción de un virión con un sitio receptor específica de la superficie celular. Las moléculas receptoras difieren para los distintos virus, y son proteínas en algunos casos (por ejemplo, picornavirus) y oligosacáridos en otros (por ejemplo, ortovirus y paramixovirus). La presencia o la ausencia de receptores desempeña una función determinante de primer orden en el tropismo celular y la patogenia viral; por ejemplo, el poliovirus es capaz de fijarse sólo en las células del sistema nervioso central y las vías digestivas de los primates. Se cree que la fijación en receptores refleja homologías configurativas fortuitas entre la estructura de la superficie del virión y un componente de la superficie celular. Por ejemplo, el virus de la inmunodeficiencia humana (HIV) se une al receptor CD4 en las células del sistema inmunitario y se cree que el virus de la rabia interactúa con los receptores de acetilcolina y que el virus de Epstein-Barr reconoce al receptor para el tercer componente del complemento en las células B. Es probable que cada célula susceptible posea por lo menos 100,000 sitios receptores para un virus dado.
Slide 3-4, Esta etapa se conoce como penetración, viropexia o fagocitosis. En algunos sistemas, se realiza por endocitosis mediada por receptores, con captación de las partículas virales ingeridas dentro de endosomas. Están menos claros los detalles de la penetración en otros sistemas. Ocurre perdida de la cubierta de manera concomitante con la penetración o poco después de la misma. La pérdida de la cubierta, o descubrimiento, es la separación física del ácido nucleico viral (o, en algunos casos, de las nucleocápsides inteARNs) a partir de los componentes estructurales externos del virión. En este momento se pierde la infecciosidad del virus original. Los virus son los únicos agentes infecciosos en los que la disolución del agente infectante es una etapa obligatoria de la vía de la replicación. Poco después de quedar descubierto el genoma viral, se inicia la fase sintética del ciclo de replicación del virus. El aspecto esencial de la replicación viral consiste en que deben transcribirse los ARNm específicos del ácido nucleico viral para que se exprese y duplique con éxito la información genética. Una vez que se logra esta etapa, los virus recurren a los componentes de la célula para traducir el ARNm. Diversas clases de virus emplean vías diferentes para sintetizar los ARNm según la estructura del ácido nucleico viral. Algunos virus (por ejemplo, rhabdovirus, ortomixovirus y paramixovirus) poseen ARN polimerasas que sintetizan a los ARNm. Los virus ARN de este tipo se llaman virus de tira negativa (de sentido negativo), puesto que su genoma de ARN de una sola tira es complementario del ARNm, el cual se designa por acuerdo general como de tira positiva (o de sentido positivo). Durante la replicación viral se sintetizan todas las macromoléculas especificadas del virus en una secuencia muy organizada. En algunas infecciones por virus, notablemente las producidas por virus de doble tira que contienen ADN, se sintetizan proteínas virales tempranas poco después de la infección y aparecen proteínas tardías sólo durante la fase tardía del proceso infeccioso, una vez que ha ocurrido la síntesis del ADN viral. Pueden activarse o no genes tempranos cuando se elaboran los productos tardíos. En contraste, se expresa al mismo tiempo la mayor parte de la información genética de los virus que contienen ARN o toda ella. Además de estos controles temporales, existen también controles cuantitativos puesto que no se elaboran todas las proteínas virales en las mismas cantidades. Las proteínas específicas del virus pueden regular la extensión de la transcripción del genoma o la traducción del ARNm viral.
Slide 5-6, Se encuentra la variación más amplia en las estrategias de la expresión genética entre los virus que contienen ARN. Algunos viriones poseen polimerasas (ortomixovirus, reovirus); otros sistemas emplean mensajes subgenómicos generados a veces por empalme (ortomixovirus, retrovirus); por último, algunos virus sintetizan grandes precursores poliproteínicos que se someten a procesamiento y segmentación para generar los productos génicos finales (picornavirus, retrovirus). La extensión a la que las enzimas específicas del virus participan en estos procesos varía de un grupo a otro. En general, los virus de mayor tamaño (herpesvirus, poxvirus) son más independientes de las funciones celulares que los virus de menor tamaño. Esta es una razón de que los virus de mayor tamaño sean más sensibles a la quimioterapia antiviral, porque se dispone de más procesos específicos de los virus como blancos de la acción farmacológica. Los sitios intracelulares en los que ocurren los diferentes acontecimientos de la replicación viral, varían de un grupo a otro. Es posible plantear algunas generalizaciones. La proteína viral se sintetiza en el citoplasma, sobre los polirribosomas compuestos por ARNm específico del virus y ribosomas de la célula huésped. El ADN viral suele replicarse en el núcleo. El ARN genómico viral se duplica generalmente en el citoplasma celular, aunque hay excepciones.
Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada La única característica de la multiplicación viral consiste en que, poco después de su interacción con una célula huésped, el virión infectante se desintegra y pierde la infecciosidad que se puede medir. Esta fase del ciclo celular se llama periodo de eclipse; su duración varía dependiendo del virus y de la célula huésped en particular, y termina con la formación de las primeras partículas virales descendientes infecciosas. El periodo de eclipse es, en realidad, de intensa actividad sintética, puesto que la célula se redirige hacia la satisfacción de las necesidades del "pirata" viral. En algunos casos, tan pronto como el ácido nucleico viral penetra a la célula huésped, el metabolismo celular es recanalizado exclusivamente hacia la síntesis de nuevas partículas virales. En otros casos, los procesos metabólicos de la célula huésped no se alteran significativamente, aunque la célula sintetiza proteínas virales y ácidos nucleicos.
La primera etapa de la infección viral es la interacción de un virión con un sitio receptor específica de la superficie celular. Las moléculas receptoras difieren para los distintos virus, y son proteínas en algunos casos (por ejemplo, picornavirus) y oligosacáridos en otros (por ejemplo, ortovirus y paramixovirus). La presencia o la ausencia de receptores desempeña una función determinante de primer orden en el tropismo celular y la patogenia viral; por ejemplo, el poliovirus es capaz de fijarse sólo en las células del sistema nervioso central y las vías digestivas de los primates. Se cree que la fijación en receptores refleja homologías configurativas fortuitas entre la estructura de la superficie del virión y un componente de la superficie celular. Por ejemplo, el virus de la inmunodeficiencia humana (HIV) se une al receptor CD4 en las células del sistema inmunitario y se cree que el virus de la rabia interactúa con los receptores de acetilcolina y que el virus de Epstein-Barr reconoce al receptor para el tercer componente del complemento en las células B. Es probable que cada célula susceptible posea por lo menos 100,000 sitios receptores para un virus dado. Después de la fijación, la partícula viral entra en la célula. Esta etapa se conoce como penetración, viropexia o fagocitosis. En algunos sistemas, se realiza por endocitosis mediada por receptores, con captación de las partículas virales ingeridas dentro de endosomas. Están menos claros los detalles de la penetración en otros sistemas. Ocurre perdida de la cubierta de manera concomitante con la penetración o poco después de la misma. La pérdida de la cubierta, o descubrimiento, es la separación física del ácido nucleico viral (o, en algunos casos, de las nucleocápsides inteARNs) a partir de los componentes estructurales externos del virión. En este momento se pierde la infecciosidad del virus original.
Los virus son los únicos agentes infecciosos en los que la disolución del agente infectante es una etapa obligatoria de la vía de la replicación Poco después de quedar descubierto el genoma viral, se inicia la fase sintética del ciclo de replicación del virus. El aspecto esencial de la replicación viral consiste en que deben transcribirse los ARNm específicos del ácido nucleico viral para que se exprese y duplique con éxito la información genética. Una vez que se logra esta etapa, los virus recurren a los componentes de la célula para traducir el ARNm. Diversas clases de virus emplean vías diferentes para sintetizar los ARNm según la estructura del ácido nucleico viral. Algunos virus (por ejemplo, rhabdovirus, ortomixovirus y paramixovirus) poseen ARN polimerasas que sintetizan a los ARNm. Los virus ARN de este tipo se llaman virus de tira negativa (de sentido negativo), puesto que su genoma de ARN de una sola tira es complementario del ARNm, el cual se designa por acuerdo general como de tira positiva (o de sentido positivo).
Durante la replicación viral se sintetizan todas las macromoléculas especificadas del virus en una secuencia muy organizada. En algunas infecciones por virus, notablemente las producidas por virus de doble tira que contienen ADN, se sintetizan proteínas virales tempranas poco después de la infección y aparecen proteínas tardías sólo durante la fase tardía del proceso infeccioso, una vez que ha ocurrido la síntesis del ADN viral. Pueden activarse o no genes tempranos cuando se elaboran los productos tardíos. En contraste, se expresa al mismo tiempo la mayor parte de la información genética de los virus que contienen ARN o toda ella. Además de estos controles temporales, existen también controles cuantitativos puesto que no se elaboran todas las proteínas virales en las mismas cantidades. Las proteínas específicas del virus pueden regular la extensión de la transcripción del genoma o la traducción del ARNm viral.
Se encuentra la variación más amplia en las estrategias de la expresión genética entre los virus que contienen ARN. Algunos viriones poseen polimerasas (ortomixovirus, reovirus); otros sistemas emplean mensajes subgenómicos generados a veces por empalme (ortomixovirus, retrovirus); por último, algunos virus sintetizan grandes precursores poliproteínicos que se someten a procesamiento y segmentación para generar los productos génicos finales (picornavirus, retrovirus).
La extensión a la que las enzimas específicas del virus participan en estos procesos varía de un grupo a otro. En general, los virus de mayor tamaño (herpesvirus, poxvirus) son más independientes de las funciones celulares que los virus de menor tamaño. Esta es una razón de que los virus de mayor tamaño sean más sensibles a la quimioterapia antiviral, porque se dispone de más procesos específicos de los virus como blancos de la acción farmacológica.
Los sitios intracelulares en los que ocurren los diferentes acontecimientos de la replicación viral, varían de un grupo a otro. Es posible plantear algunas generalizaciones. La proteína viral se sintetiza en el citoplasma, sobre los polirribosomas compuestos por ARNm específico del virus y ribosomas de la célula huésped. El ADN viral suele replicarse en el núcleo. El ARN genómico viral se duplica generalmente en el citoplasma celular, aunque hay excepciones.
El ciclo de multiplicación viral se define como la serie de etapas secuenciales que transcurren desde el contacto inicial de un virus con una célula huésped hasta la liberación a partir de la misma célula infectada, de la progenie de nuevos viriones.
El primer paso en la infección de una célula es la adhesión a la superficie celular. Se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos en el exterior de la célula. Las células que carecen de los receptores apropiados no son susceptibles al virus.
Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus.
(A) Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular.
(B) Entada vía endosomas en la superficie celular
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
3. PÉRDIDA DE LA CÁPSULA El ácido nucleico debe de estar lo suficientemente expuesto como para que la replicación viral comience en esta fase. Cuando el ácido nucleico es expuesto, las partículas víricas infecciosas no pueden removerse de las células – este es el comienzo de la fase de ECLIPSE – la cual perdura hasta que viriones infecciosos nuevos sean creados.
4. SÍNTESIS DE ÁCIDO NUCLEICO Y PROTEINAS VIRALES Existen diversos mecanismos, algunos serán abarcados en capítulos posteriores.
5. ENSAMBLAJE/MADURACIÓN Nuevas partículas víricas son ensambladas. Puede haber un estadio de maduración posterior al proceso inicial de ensamblaje.
6. LIBERACIÓN O DESCARGA Los virus pueden ser liberados mediante lisis celular, o, si son envueltos, pueden yemar de las células. Los virus que yeman no necesariamente destruyen la célula. Por tanto, algunos de estos virus son capaces de instaurar infecciones persistentes. No todas las partículas víricas liberadas son infecciosas. La proporción partículas infecciosas:partículas no-infecciosas varía según el virus y las condiciones de crecimiento.
#2012-1270 Adhesión El evento inicial en el ciclo de vida de un virus es la unión del mismo con la célula huésped a través del proceso de adsorción. Esta unión específica se establece entre una molécula presente en la membrana celular (receptor) y una proteína externa del virión. Algunos receptores virales han sido identificados y son en general carbohidratos, lípidos o proteínas de la membrana plasmática.
Así, la susceptibilidad o resistencia de una célula a la infección está determinada por la presencia o ausencia de receptores específicos. La importancia de la carencia de receptores se demuestra indirectamente porque es posible infectar exitosamente células no susceptibles a algunos virus utilizando ácido nucleico desnudo (transfección).
La unión virus-célula ocurre en dos etapas: una primera unión reversible por atracción electrostática, facilitada por la presencia de cationes: la adsorción irreversible se produce cuando se establece la unión polivalente virus-célula, ya que suele haber muchas copias de los receptores en la célula y múltiple sitios de unión en el virión.
#2012-1270 Penetración Una vez establecida la adsorción, los virus pueden entrar en la célula a través de tres mecanismos diferentes: endocitosis mediada por receptor, fusión con la membrana plasmática y traslocación a través de la membrana plasmática.
Se acepta en la actualidad que los virus desnudos pueden entrar en la célula por traslocación o por endocitosis mientras que los virus envueltos penetran por endocitosis o por fusión.
En la endocitosis mediada por receptor el virus utiliza la misma vía de entrada que emplean muchas macromoléculas fisiológicamente importantes como nutrientes hormonas y factores de crecimiento. El virus absorbido a su receptor va quedando encerrado dentro de invaginaciones de la membrana plasmática recubiertas de una proteína llamada clatrina, que forman primero un hueco recubierto y luego una vesícula. De este modo el virión intacto entra en el endosoma. El pH ácido del endosoma (5 a 5,5) produce cambios conformacionales en las proteínas del virión que permiten la fusión de la membrana del endosoma con la membrana externa del virus y expulsan el genoma viral al citoplasma celular para su libre expresión.
El mecanismo de fusión directa de la envoltura viral con la membrana plasmática celular es utilizado por ciertos virus envueltos como herpes y Paramixovirus. Estos virus poseen en su envoltura glicoproteínas con capacidad de fusión independiente del pH, es decir, que a un pH neutro ocurre la fusión a nivel de la membrana celular liberando el genoma al citoplasma. A diferencia de la entrada por endocitosis, en este caso en ningún momento se ven viriones intactos dentro de la célula. Esta capacidad de fusión con membrana plasmática que tienen ciertos virus ha permitido su utilización como agentes fusogénicos para la obtención de híbridos celulares. (Ver virus del VIH)
Por último, algunos virus desnudos como Adenovirus parecen entrar directamente a través de la membrana plasmática mediante un proceso de traslocación.
El ciclo de replicación de un bacteriófago se puede dividir esquemáticamente en distintas etapas, las que son comunes a otros virus bacterianos y eucarióticos.
Adsorción: El virus se fija o adsorbe a componentes de la superficie celular que actúan como receptores específicos. La zona de adsorción del virus es complementaria al receptor celular, por lo tanto un determinado virus sólo puede infectar un número limitado de cepas celulares que contengan a un determinado receptor. La naturaleza de la zona de adsorción varía con el tipo de fago. En el T4 se localiza en el extremo de la cola, en donde se encuentran la placa basal, las espículas y las fibras de la cola.
Inyección del material genético viral: Después de la adsorción, se produce un cambio configuracional en las proteínas de la placa basal. Mediante su actividad enzimática producen un poro en la membrana citoplasmática de la célula. La vaina del fago se contrae y el material genético viral ingresa en la célula, mientras que la envoltura proteica queda en el exterior.
Replicación del material genético viral: El material genético viral que ingresa en una célula contiene bases modificadas que evitan la degradación por nucleasas bacterianas. Esta modificación consiste en la glicosilación y/o metilación de algunas determinadas bases. En el caso del fago T4 se glucosila la base 5'-hidroximetilcitosina. Se deben sintetizar proteínas tempranas que reparan el poro de la membrana citoplasmática por donde ingresó el genoma viral, degradan el DNA bacteriano lo que proporciona una fuente de precursores, evita la síntesis de RNA y proteínas bacterianas, y proporciona ribosomas para la síntesis de proteínas del fago. Además algunas de estas proteínas tempranas participan en la síntesis de las bases inusuales. La forma de replicación del genoma viral es dependiente del tipo de material genético (si es RNA o DNA, si es simple o doble cadena).
Síntesis de las envolturas proteicas: Las proteínas de la envoltura son proteínas tardías que se sintetizan después de iniciada la replicación del material genético.
Ensamble: Todas las proteínas de la envoltura se ensamblan para formar una partícula viral madura capaz de infectar a otra célula cuando sea liberada.
Lisis celular y liberación de las partículas virales: La lisis celular se debe a la síntesis de proteínas tardías codificadas en el genoma del fago.
#2012-1270 Replicación y maduración REPLICACION DE VIRUS DNA Después de la adsorción, penetración y eliminación de la cubierta, el ácido nucleico viral es liberado y migra al núcleo de la célula hospedadora. Se transcribe la porción de DNA de los genes tempranos. Tiene lugar la traducción, siendo los productos de estos genes enzimas requeridos para la multiplicación del DNA vírico. En la mayoría de los virus DNA, la transcripción temprana se lleva a cabo con la transcriptasa (RNA polimerasa) del hospedador. Tiempo después comienza la transcripción y traducción de los genes tardíos. La síntesis de las proteínas de la cápsida ocurre en el citoplasma de la célula hospedadora. Las proteínas de la cápsida migran al núcleo donde tiene lugar la maduración: el DNA vírico y las proteínas de la cápsida se ensamblan para formar las partículas víricas completas. REPLICACION DE VIRUS RNA a) VIRUS DE ARN QUE NO POSEEN UNA FASE DE ADN Los virus que se replican vía intermediarios de ARN necesitan una ARN – polimerasa dependiente de ARN para replicar su ARN, pero las células animales aparentemente no tienen esa enzima. Por tanto, este tipo de virus de ARN necesita codificar su ARN – polimerasa dependiente de ARN. Ninguna proteína viral puede ser sintetizada hasta que el ARN mensajero del virus esté disponible. i. Virus de ARN de cadena positiva En estos virus, el ARN del virión tiene el mismo sentido que un ARNm y funciona como tal. Este ARNm puede ser traducido inmediatamente luego de la infección. ii. Virus de ARN de cadena negativa El ARN del virión tiene sentido negativo (complementario al ARNm). Se copia en un ARNm de sentido positivo complementario antes de que las proteínas puedan ser sintetizadas. Entonces, necesitan codificar para una ARN – polimerasa dependiente de ARN y empacarla en el virión para sintetizar ARNm cuando infectan. iii. Virus de ARN de cadena doble No puede funcionar como ARNm; necesitan también empacar una ARN – polimerasa para sintetizar su ARNm luego de producir infecciones en células huésped. b) VIRUS DE ARN QUE COPIAN SU ARN EN ADN. Son los retrovirus de ARN sentido positivo que no funciona como ARNm luego de la infección puesto que no es liberado de su cápside al citoplasma. Sirve de plantilla para la transcriptasa inversa y es copiado en ADN. La enzima no está disponible en la célula se codifica y empaca en los viriones.
REPLICACION DE VIRUS DNA Después de la adsorción, penetración y eliminación de la cubierta, el ácido nucleico viral es liberado y migra al núcleo de la célula hospedadora. Se transcribe la porción de DNA de los genes tempranos. Tiene lugar la traducción, siendo los productos de estos genes enzimas requeridos para la multiplicación del DNA vírico. En la mayoría de los virus DNA, la transcripción temprana se lleva a cabo con la transcriptasa (RNA polimerasa) del hospedador. Tiempo después comienza la transcripción y traducción de los genes tardíos. La síntesis de las proteínas de la cápsida ocurre en el citoplasma de la célula hospedadora. Las proteínas de la cápsida migran al núcleo donde tiene lugar la maduración: el DNA vírico y las proteínas de la cápsida se ensamblan para formar las partículas víricas completas. REPLICACION DE VIRUS RNA a) VIRUS DE ARN QUE NO POSEEN UNA FASE DE ADN Los virus que se replican vía intermediarios de ARN necesitan una ARN – polimerasa dependiente de ARN para replicar su ARN, pero las células animales aparentemente no tienen esa enzima. Por tanto, este tipo de virus de ARN necesita codificar su ARN – polimerasa dependiente de ARN. Ninguna proteína viral puede ser sintetizada hasta que el ARN mensajero del virus esté disponible. i. Virus de ARN de cadena positiva En estos virus, el ARN del virión tiene el mismo sentido que un ARNm y funciona como tal. Este ARNm puede ser traducido inmediatamente luego de la infección. ii. Virus de ARN de cadena negativa El ARN del virión tiene sentido negativo (complementario al ARNm). Se copia en un ARNm de sentido positivo complementario antes de que las proteínas puedan ser sintetizadas. Entonces, necesitan codificar para una ARN – polimerasa dependiente de ARN y empacarla en el virión para sintetizar ARNm cuando infectan. iii. Virus de ARN de cadena doble No puede funcionar como ARNm; necesitan también empacar una ARN – polimerasa para sintetizar su ARNm luego de producir infecciones en células huésped. b) VIRUS DE ARN QUE COPIAN SU ARN EN ADN. Son los retrovirus de ARN sentido positivo que no funciona como ARNm luego de la infección puesto que no es liberado de su cápside al citoplasma. Sirve de plantilla para la transcriptasa inversa y es copiado en ADN. La enzima no está disponible en la célula se codifica y empaca en los viriones.
#2012-1270 Maduración y salida de virus desnudos y envueltos El evento inicial en el ciclo de vida de un virus es la unión del mismo con la célula huésped a través de un receptor, proceso de adsorción. En la endocitosis mediada por receptor, el virus va quedando encerrado dentro de invaginaciones de la membrana. De este modo el virión intacto entra en el endosoma. En cambio, el mecanismo de fusión directa de la envoltura viral con la membrana plasmática celular es utilizado por ciertos virus envueltos que poseen glicoproteínas con capacidad de fusión independiente del pH. El desnudamiento ocurre simultáneamente o después de la penetración y comprende la eliminación de todas o parte de las cubiertas proteicas que rodean al ácido nucleico viral, de manera de dejar expuesto dentro de la célula el genoma viral desnudo o asociado a alguna enzima para que pueda expresarse. Se sintetizan proteínas virales y se replica el ácido nucleico viral. Virus con genoma de ADN: Los virus llevan a cabo la duplicación de su ADN por medio de mecanismos y herramientas similares a los empleados por la célula para replicar su material genético. La replicación del ADN viral es generalmente semiconservativa, es decir cada cadena del ADN parental se conserva y por apareamiento con las bases complementarias se duplica, produciéndose como resultado final dos moléculas de ADN, cada una de las cuales está formada por una cadena de ADN parental y una cadena sintetizada "de novo". Virus con genoma de ARN: A diferencia de lo que sucede con el ADN, la replicación del ARN es un fenómeno único de los virus ya que la maquinaria genética de la célula se basa exclusivamente en el uso del ADN como templado (ADN-ADN o ADN-ARN). Por lo tanto, para la duplicación de su genoma, así como acontecía para la síntesis de sus ARNm, los virus ARN deben necesariamente aportar la enzima ARN polimerasa ARN dependiente. En las etapas tardías del ciclo de multiplicación viral las moléculas de ácido nucleico y proteínas virales sintetizadas en la célula infectada deben ensamblarse para formar los nuevos viriones y salir al espacio extracelular para estar en condiciones de infectar nuevas células. Este proceso de maduración y ensamble, también denominado morfogénesis, comprende el armado de la cápside a partir de las unidades estructurales y luego la formación del virión completo. El mecanismo va a diferir en base a dos propiedades del virión: la estructura de la cápside y la presencia o no de envoltura.
#2012-1270 Salida de un virus envuelto Los virus envueltos en el último paso al ensamblaje, el virion, se une con la membrana celular a la salida de la célula, mediante proteínas que se insertan en la superficie interna y externa de la membrana. El virion naciente brota a través de la membrana. El ensamblaje y maduración con la expulsión a través de la superficie celular proporcionan un mecanismo más eficaz de salida, ya que no depende de la desintegración de la célula infectada. A la inserción de glicoproteínas virales en la superficie celular se cambia la especifidad antigénica, con lo que la célula infectada se convierte en blanco para el aparato inmunológico del huésped.
Wladimir A. Rijo Inirio 89257 D# 1(Etapas de la replicacion Viral)
Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada.
El primer paso en la infección de una célula es la adhesión o fijación a la superficie celular. Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula. Las células que carecen de los receptores apropiados no son susceptibles al virus.
Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus. Virus envueltos (A) Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular. (B) Entada vía endosomas en la superficie celular. Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula. Virus no envueltos o desnudos Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
Wladimir A. Rijo Inirio 89257 D# 4(Penetracion de los Bacteriografos)
Los virus complejos producen una rotura en la membrana celular del hospedador en uno de los puntos de anclaje, gracias a la presencia de algunas moléculas de enzimas hidrolíticas entre las proteínas de la cápside. A través de la rotura, el tubo central inyecta el ADN vírico, quedando la cápside vacía en el exterior de la célula diana y el ácido nucleico libre en el citoplasma. La presencia de cápsidas en la superficie celular es un buen indicio de que ha sufrido una infección vírica. Otros virus sin envoltura lipídica se introducen en la célula con cápsida y todo, lo cual puede realizarse de dos maneras: Por penetración directa: después de la fijación, el virus abre una brecha en la membrana y se introduce en el citoplasma. Por endocitosis: la membrana forma una invaginación en torno al virus, llegando a formar una vesícula que penetra en la célula. Formada la vesícula, el virus abre una brecha en la membrana de la misma con ayuda de algunas enzimas hidrolíticas que él mismo transporta, penetrando así en el citoplasma.
Los virus con envoltura lipídica burlan la barrera de la membrana celular porque su cubierta lipídica se funde con la membrana, ya que son de la misma naturaleza. Esta fusión de membranas puede realizarse en dos lugares distintos: Fusión en la superficie celular: de manera que el virión penetra directamente en el citoplasma.
Fusión con un lisosoma: se forma una vesícula por endocitosis, a la que se une un lisosoma para digerir la partícula introducida; entonces, la cubierta lipídica del virus se funde con la membrana del lisosoma y el virión escapa hacia el citoplasma.
Wladimir A. Rijo Inirio 89257 D# 5 (Replicacion y Maduracion)
Ensamblaje de los componentes de los virus para formar nuevas partículas víricas (proceso también llamado maduración). Los virus desnudos realizan su ensamblaje y maduración en forma intracelular y adquieren capacidad infectante dentro de la célula. Para su salida requieren de la desintegración celular. Los virus envueltos en el último paso al ensamblaje el virion se une con la membrana celular a la salida de la célula, mediante proteínas que se insertan en la superficie interna y exterior de la membrana. El virion naciente brota atraves de la membrana. El ensamblaje y maduración con la expulsión atraves de la superficie celular proporcionan un mecanismo más eficaz de salida, ya que no depende de la desintegración de la célula infectada.
Wladimir A. Rijo Inirio 89257 D# 6(Maduracion y salida de virus desnudos y envueltos)
Después de la multiplicación del virus tiene lugar la salida de los nuevos individuos, que saldrán con capacidad de infectar nuevas células. Las principales modalidades de liberación da nombre a nuevas variantes del ciclo vital de los virus:
Infección persistente Los nuevos virus no esperan a la muerte de la célula hospedadora para abandonarla, sino que van saliendo de la célula al mismo tiempo que se van produciendo, de manera que la célula puede seguir viva y produciendo nuevas partículas víricas. La liberación puede hacerse de dos maneras: Los virus sin envoltura lipoproteica salen directamente, sin arrastrar ningún resto de la membrana plasmática, bien sea abriendo una brecha en la membrana, o bien aprovechando los mecanismos de exocitosis o salida de sustancias al exterior de la célula.
Wladimir A. Rijo Inirio 89257 D# 7(Salida de virus envuelto)
Liberación de los nuevos virus de la célula hospedadora, que ya pueden infectar otras células. Pueden producirse con o sin rotura de la célula, dependiendo de los virus. Los virus desnudos salen por lisis celular lo contrario de los envueltos que salen por gemación.
88032 Slide # 2 y3 Adhesion y Penetracion El primer paso en la infección de una célula es la adhesión o fijación a la superficie celular. Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula. Las células que carecen de los receptores apropiados no son susceptibles al virus. La penetración: Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus.Virus envueltos Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular. Entada vía endosomas en la superficie celular. Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula. Virus no envueltos o desnudos Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
88032 slide #4 La primera etapa de la infección viral es la interacción de un virión con un sitio receptor específica de la superficie celular. Las moléculas receptoras difieren para los distintos virus, y son proteínas en algunos casos (por ejemplo, picornavirus) y oligosacáridos en otros (por ejemplo, ortovirus y paramixovirus). La presencia o la ausencia de receptores desempeña una función determinante de primer orden en el tropismo celular y la patogenia viral; por ejemplo, el poliovirus es capaz de fijarse sólo en las células del sistema nervioso central y las vías digestivas de los primates. Se cree que la fijación en receptores refleja homologías configurativas fortuitas entre la estructura de la superficie del virión y un componente de la superficie celular. El virus de la inmunodeficiencia humana (HIV) se une al receptor CD4 en las células del sistema inmunitario y se cree que el virus de la rabia interactúa con los receptores de acetilcolina y que el virus de Epstein-Barr reconoce al receptor para el tercer componente del complemento en las células B. Es probable que cada célula susceptible posea por lo menos 100,000 sitios receptores para un virus dado
88032 Slide #5 Replicacion y Maduracion Ensamblaje de los componentes de los virus para formar nuevas partículas víricas. Los virus desnudos realizan su ensamblaje y maduración en forma intracelular y adquieren capacidad infectante dentro de la célula. Para su salida requieren de la desintegración celular. Los virus envueltos en el último paso al ensamblaje el virion se une con la membrana celular a la salida de la célula, mediante proteínas que se insertan en la superficie interna y exterior de la membrana. El virion naciente brota atraves de la membrana. El ensamblaje y maduración con la expulsión atraves de la superficie celular proporcionan un mecanismo más eficaz de salida, ya que no depende de la desintegración de la célula infectada.
88032 Slide #6 Maduracion y Lberacion Los virus en general pueden liberarse de la célula mediante dos tipos de mecanismos: lisis y gemación. Para los virus líticos -la mayoría de los virus desnudos-, la liberación es un proceso simple: la célula infectada se rompe y libera las partículas virales. En el caso de los virus que se liberan por gemación, estos adquieren su envoltura lipídica y las proteínas asociadas en el momento en que pasan a través de la membrana plasmática o de vesículas intracelulares. La liberación de los virus por esta vía puede resultar altamente perjudicial para la célula -por ej.en el caso de paramixovirus, rabdovirus, togavirus- o no -por ej. retrovirus-; de todos modos en cualquiera de los casos el proceso es controlado por el virus: la interacción física de las proteínas de la cápside sobre la superficie interna de la membrana celular fuerza a la partícula viral a pasar a través de dicha membrana. El ensamble, la maduración y liberación son generalmente procesos simultáneos en los virus que se liberan por gemación. El tipo de membrana a través de la cual geman los virus depende del tipo de virus, por ej. puede ser a través de la membrana celular como en el caso de los retrovirus y togavirus, a través de la membrana citoplasmática -ortomixovirus, coronavirus, bunyavirus, rabdovirus, hepadnavirus- o a través de la membrana nuclear como en los herpesvirus. Slide 7: Salida de un virus envuelto 2012-0593 Se encuentra la variación más amplia en las estrategias de la expresión genética entre los virus que contienen ARN. Algunos viriones poseen polimerasas (ortomixovirus, reovirus); otros sistemas emplean mensajes subgenómicos generados a veces por empalme (ortomixovirus, retrovirus); por último, algunos virus sintetizan grandes precursores poliproteínicos que se someten a procesamiento y segmentación para generar los productos génicos finales (picornavirus, retrovirus).Los sitios intracelulares en los que ocurren los diferentes acontecimientos de la replicación viral, varían de un grupo a otro. Es posible plantear algunas generalizaciones. La proteína viral se sintetiza en el citoplasma, sobre los polirribosomas compuestos por ARNm específico del virus y ribosomas de la célula huésped. El ADN viral suele replicarse en el núcleo.
88032 Slide #7 Salida del virus envuelto Liberación de los nuevos virus de la célula hospedadora, que ya pueden infectar otras células. Pueden producirse con o sin rotura de la célula, dependiendo de los virus. Los virus desnudos salen por lisis celular lo contrario de los envueltos que salen por gemación. -todo Se encuentra la variación más amplia en las estrategias de la expresión genética entre los virus que contienen ARN. Algunos viriones poseen polimerasas (ortomixovirus, reovirus); otros sistemas emplean mensajes subgenómicos generados a veces por empalme (ortomixovirus, retrovirus). Dependiendo del virus tiene distintos manera de salida virus desnudos salen por lisis celular y los envueltos que salen por gemación.
1.Etapas de la replicacion de virosis El Ciclo De Replicación De Los Virus consta generalmente de las siguientes fases: fijación y entrada en la célula, eclipse, multiplicación y liberación del virus.
El primer paso en la infección viral es la absorción a la membrana de la célula susceptible por medio de la adhesión de ligandos virales (proteínas de la capside o glucoproteínas de las espículas, por ejemplo a receptores superficiales de la célula). La distribución en el cuerpo del hospedador de ciertos tipos de receptores celulares explica el tropismo tisular y de hospedador de los virus. Los tejidos y células que carecen de receptores específicos de virus determinados no son infectados por dichos viriones. La naturaleza de los receptores es variable y ciertos virus pueden tener más de un tipo de receptor.
3.Penetracion Los virus complejos producen una rotura en la membrana celular del hospedador en uno de los puntos de anclaje, gracias a la presencia de algunas moléculas de enzimas hidrolíticas entre las proteínas de la cápsida. A través de la rotura, el tubo central inyecta el ADN vírico, quedando la cápside vacía en el exterior de la célula diana y el ácido nucleico libre en el citoplasma. La presencia de cápsidas en la superficie celular es un buen indicio de que ha sufrido una infección vírica. Otros virus sin envoltura lipídica se introducen en la célula con cápsida y todo, lo cual puede realizarse de dos maneras: Por penetración directa: después de la fijación, el virus abre una brecha en la membrana y se introduce en el citoplasma. Por endocitosis: la membrana forma una invaginación en torno al virus, llegando a formar una vesícula que penetra en la célula. Formada la vesícula, el virus abre una brecha en la membrana de la misma con ayuda de algunas enzimas hidrolíticas que él mismo transporta, penetrando así en el citoplasma.
4.Penetracion de los bacteriofagos Adsorción: El virus se fija o adsorbe a componentes de la superficie celular que actúan como receptores específicos. La zona de adsorción del virus es complementaria al receptor celular, por lo tanto un determinado virus sólo puede infectar un número limitado de cepas celulares que contengan a un determinado receptor. La naturaleza de la zona de adsorción varía con el tipo de fago. En el T4 se localiza en el extremo de la cola, en donde se encuentran la placa basal, las espículas y las fibras de la cola. Inyección del material genético viral: Después de la adsorción, se produce un cambio configuracional en las proteínas de la placa basal, alguna de las cuales tienen actividad enzimática y producen un poro en la membrana citoplasmática de la célula. La vaina del fago se contrae y el material genético viral ingresa en la célula, mientras que la envoltura proteica queda en el exterior. Replicación del material genético viral: El material genético viral que ingresa en una célula contiene bases modificadas que evitan la degradación por nucleasas bacterianas. Esta modificación consiste en la glicosilación y/o metilación de algunas determinadas bases. En el caso del fago T4 se glucosila la base 5'-hidroximetilcitosina. Para lograr una efectiva replicación del genoma viral se deben sintetizar algunas proteínas ni bien el material genético ingresa en la célula. Síntesis de las envolturas proteicas: Las proteínas de la envoltura (cápside, vaina, fibras, etc) son proteínas tardías que se sintetizan después de iniciada la replicación del material genético. La síntesis de cada componente proteico se realiza separadamente. En el caso del fago T4, el material genético es encapsidado antes del ensamble del resto de los componentes. Ensamble: Todas las proteínas de la envoltura se ensamblan para formar una partícula viral madura capaz de infectar a otra célula cuando sea liberada. Lisis celular y liberación de las partículas virales: La lisis celular se debe a la síntesis de proteínas tardías codificadas en el genoma del fago. En el fago T4, estas proteínas son enzimas que lesionan la membrana citoplasmática y la pared celular.
5.Replicacion y Maduracion La multiplicación del virus consta de la replicación de su material genético, de la transcripción de su mensaje en una molécula de ARN y de la traducción del mensaje para producir proteínas víricas, tanto las que formarán parte de la cápsida como las proteínas enzimáticas necesarias para el ensamblaje de las piezas del virión y para algunas de las funciones anteriores. Los ribosomas y la mayor parte de las enzimas que los ácidos nucleicos víricos utilizan en estos procesos son los de la célula infectada. Posteriormente a estos procesos, tiene lugar el ensamblaje de las piezas para construir nuevos viriones. En muchos virus, como el del mosaico del tabaco, el ensamblaje es automático y depende de la concentración salina del medio. En otros virus, en el ensamblaje intervienen enzimas codificadas en el ácido nucleico del virus.
6.Maduracion y salida de virus envueltos y desnudos Los virus desnudos realizan su ensamble y maduracion en forma intracelular, y adquieren capacidad infectante dentro de la celula. Para su salida requieren de la desintegracion celular. La mayoría de los virus desnudos son resistentes al medio externo, a la desecación y a los solventes de lípidos. Por esta razón, los virus desnudos pueden penetrar por vía digestiva ya que resisten el pH ácido del estómago. Pueden ser transmitidos por vía fecal.oral, a través de agua o alimentos contaminados o por manos u objetos. Son ejemplos de virus desnudos el Poliovirus, y los Adenovirus. Los virus envueltos en el ultimo paso al ensamble, se unen con la membrana celular a la salida de la celula, mediante proteinas que se insertan en la superficie interna y exterior de la membrana.
7.Salida de un virus envuelto Los virus envueltos en el ultimo paso al ensamble (el virion), se unen con la membrana celular a la salida de la celula, mediante proteinas que se insertan en la superficie interna y exterior de la membrana. Al final se obtiene un virion infeccioso con envoltura de glicoproteinas.
87451 Felix Bienvenido Carrasco Vasquez grupo: lunes 10-12
Adhesión
La adhesión es la propiedad de la materia por la cual se unen y plasman dos superficies de sustancias iguales o diferentes cuando entran en contacto, y se mantienen juntas por fuerzas intermoleculares. La adhesión ha jugado un papel muy importante en muchos aspectos de las técnicas de construcción tradicionales. La adhesión del ladrillo con el mortero (cemento) es un ejemplo claro. La cohesión es distinta de la adhesión. La cohesión es la fuerza de atracción entre partículas adyacentes dentro de un mismo cuerpo, mientras que la adhesión es la interacción entre las superficies de distintos cuerpos.
87451 Felix Bienvenido Carrasco Vasquez grupo: lunes 10-12
Penetración
Los virus complejos producen una rotura en la membrana celular del hospedador en uno de los puntos de anclaje, gracias a la presencia de algunas moléculas de enzimas hidrolíticas entre las proteínas de la cápsida. A través de la rotura, el tubo central inyecta el ADN vírico, quedando la cápside vacía en el exterior de la célula diana y el ácido nucleico libre en el citoplasma. La presencia de cápsidas en la superficie celular es un buen indicio de que ha sufrido una infección vírica. Otros virus sin envoltura lipídica se introducen en la célula con cápsida y todo, lo cual puede realizarse de dos maneras: Por penetración directa: después de la fijación, el virus abre una brecha en la membrana y se introduce en el citoplasma. Por endocitosis: la membrana forma una invaginación en torno al virus, llegando a formar una vesícula que penetra en la célula. Formada la vesícula, el virus abre una brecha en la membrana de la misma con ayuda de algunas enzimas hidrolíticas que él mismo transporta, penetrando así en el citoplasma.
87451 Felix Bienvenido Carrasco Vasquez grupo: lunes 10-12
Penetracion de los bacteriofagos
Adsorción: El virus se fija o adsorbe a componentes de la superficie celular que actúan como receptores específicos. La zona de adsorción del virus es complementaria al receptor celular, por lo tanto un determinado virus sólo puede infectar un número limitado de cepas celulares que contengan a un determinado receptor. La naturaleza de la zona de adsorción varía con el tipo de fago. En el T4 se localiza en el extremo de la cola, en donde se encuentran la placa basal, las espículas y las fibras de la cola.
Inyección del material genético viral: Después de la adsorción, se produce un cambio configuracional en las proteínas de la placa basal, alguna de las cuales tienen actividad enzimática y producen un poro en la membrana citoplasmática de la célula. La vaina del fago se contrae y el material genético viral ingresa en la célula, mientras que la envoltura proteica queda en el exterior. Replicación del material genético viral: El material genético viral que ingresa en una célula contiene bases modificadas que evitan la degradación por nucleasas bacterianas. Esta modificación consiste en la glicosilación y/o metilación de algunas determinadas bases. En el caso del fago T4 se glucosila la base 5'- hidroximetilcitosina. Para lograr una efectiva replicación del genoma viral se deben sintetizar algunas proteínas ni bien el material genético ingresa en la célula. Síntesis de las envolturas proteicas: Las proteínas de la envoltura (cápside, vaina, fibras, etc) son proteínas tardías que se sintetizan después de iniciada la replicación del material genético. La síntesis de cada componente proteico se realiza separadamente. En el caso del fago T4, el material genético es encapsidado antes del ensamble del resto de los componentes.
Ensamble: Todas las proteínas de la envoltura se ensamblan para formar una partícula viral madura capaz de infectar a otra célula cuando sea liberada. Lisis celular y liberación de las partículas virales: La lisis celular se debe a la síntesis de proteínas tardías codificadas en el genoma del fago. En el fago T4, estas proteínas son enzimas que lesionan la membrana citoplasmática y la pared celular.
87451 Felix Bienvenido Carrasco Vasquez grupo: lunes 10-12
Replicacion y Maduracion
La multiplicación del virus consta de la replicación de su material genético, de la transcripción de su mensaje en una molécula de ARN y de la traducción del mensaje para producir proteínas víricas, tanto las que formarán parte de la cápsida como las proteínas enzimáticas necesarias para el ensamblaje de las piezas del virión y para algunas de las funciones anteriores. Los ribosomas y la mayor parte de las enzimas que los ácidos nucleicos víricos utilizan en estos procesos son los de la célula infectada. Posteriormente a estos procesos, tiene lugar el ensamblaje de las piezas para construir nuevos viriones. En muchos virus, como el del mosaico del tabaco, el ensamblaje es automático y depende de la concentración salina del medio. En otros virus, en el ensamblaje intervienen enzimas codificadas en el ácido nucleico del virus.
87451 Felix Bienvenido Carrasco Vasquez grupo: lunes 10-12
Maduracion y salida de virus envueltos y desnudos
Los virus desnudos realizan su ensamble y maduracion en forma intracelular, y adquieren capacidad infectante dentro de la celula. Para su salida requieren de la desintegracion celular. La mayoría de los virus desnudos son resistentes al medio externo, a la desecación y a los solventes de lípidos. Por esta razón, los virus desnudos pueden penetrar por vía digestiva ya que resisten el pH ácido del estómago. Pueden ser transmitidos por vía fecal.oral, a través de agua o alimentos contaminados o por manos u objetos. Son ejemplos de virus desnudos el Poliovirus, y los Adenovirus. Los virus envueltos en el ultimo paso al ensamble, se unen con la membrana celular a la salida de la celula, mediante proteinas que se insertan en la superficie interna y exterior de la membrana.
1- El primer paso en la infección viral es la absorción a la membrana de la célula susceptible por medio de la adhesión de ligandos virales (proteínas de la capside o glucoproteínas de las espículas, por ejemplo a receptores superficiales de la célula). La distribución en el cuerpo del hospedador de ciertos tipos de receptores celulares explica el tropismo tisular y de hospedador de los virus. Los tejidos y células que carecen de receptores específicos de virus determinados no son infectados por dichos viriones. La naturaleza de los receptores es variable y ciertos virus pueden tener más de un tipo de receptor. 2- Los virus complejos producen una rotura en la membrana celular del hospedador en uno de los puntos de anclaje, gracias a la presencia de algunas moléculas de enzimas hidrolíticas entre lasproteínas de la cápsida. A través de la rotura, el tubo central inyecta el ADN vírico, quedando la cápside vacía en el exterior de la célula diana y el ácido nucleico libre en el citoplasma. La presencia de cápsidas en la superficie celular es un buen indicio de que ha sufrido una infección vírica.
Otros virus sin envoltura lipídica se introducen en la célula con cápsida y todo, lo cual puede realizarse de dos maneras: • Por penetración directa: después de la fijación, el virus abre una brecha en la membrana y se introduce en el citoplasma. • Por endocitosis: la membrana forma una invaginación en torno al virus, llegando a formar una vesícula que penetra en la célula. Formada la vesícula, el virus abre una brecha en la membrana de la misma con ayuda de algunas enzimas hidrolíticas que él mismo transporta, penetrando así en el citoplasma. Los virus con envoltura lipídica burlan la barrera de la membrana celular porque su cubierta lipídica se funde con la membrana, ya que son de la misma naturaleza. Esta fusión de membranas puede realizarse en dos lugares distintos: • Fusión en la superficie celular: de manera que el virión penetra directamente en el citoplasma. • Fusión con un lisosoma: se forma una vesícula por endocitosis, a la que se une un lisosoma para digerir la partícula introducida; entonces, la cubierta lipídica del virus se funde con la membrana del lisosoma y el virión escapa hacia el citoplasma.
3- Según la duración de la fase de eclipse, se suelen distinguir dos modalidades de ciclo infeccioso de un virus: • Ciclo ordinario: el ácido nucleico vírico procede inmediatamente a la transcripción de su mensaje genético en los ARN necesarios para su multiplicación, y prosigue rápidamente el ciclo vital. Este tipo de ciclo es el más extendido en la naturaleza. • Ciclo lisogénico: fue descubierto por André Lwoff en bacteriófagos. El ADN vírico se cierra por sus extremos generando un ADN circular. Este ADN se inserta en el ADN bacteriano en un lugar específico en el que la secuencia de nucleótidos bacterianos es semejante alguna región del ADN vírico.
Algunos virus que infectan células animales siguen también el ciclo lisogénico, como los papilomavirus de las verrugas y algunos retrovirus que producen algunos tipos de cáncer. En el caso de los retrovirus, conviene recordar que el ácido nucleico es ARN monocatenario, por lo que la transcriptasa inversa ha de copiar el genoma vírico en forma de ADN antes de que pueda insertarse en el ADN celular. 4- La multiplicación del virus consta de la replicación de su material genético, de la transcripción de su mensaje en una molécula de ARN y de la traducción del mensaje para producir proteínasvíricas, tanto las que formarán parte de la cápsida como las proteínas enzimáticas necesarias para el ensamblaje de las piezas del virión y para algunas de las funciones anteriores. Losribosomas y la mayor parte de las enzimas que los ácidos nucleicos víricos utilizan en estos procesos son los de la célula infectada.
Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada.
La única característica de la multiplicación viral consiste en que, poco después de su interacción con una célula huésped, el virión infectante se desintegra y pierde la infecciosidad que se puede medir. Esta fase del ciclo celular se llama periodo de eclipse; su duración varía dependiendo del virus y de la célula huésped en particular, y termina con la formación de las primeras partículas virales descendientes infecciosas. El periodo de eclipse es, en realidad, de intensa actividad sintética, puesto que la célula se redirige hacia la satisfacción de las necesidades del "pirata" viral. En algunos casos, tan pronto como el ácido nucleico viral penetra a la célula huésped, el metabolismo celular es recanalizado exclusivamente hacia la síntesis de nuevas partículas virales. En otros casos, los procesos metabólicos de la célula huésped no se alteran significativamente, aunque la célula sintetiza proteínas virales y ácidos nucleicos.
Los virus han desarrollado diversas estrategias para lograr la multiplicación en las células huéspedes parasitadas por ellos. Aunque los detalles varían de un grupo a otro, el patrón general de los ciclos de replicación es semejante. La primera etapa de la infección viral es la interacción de un virión con un sitio receptor específica de la superficie celular. Las moléculas receptoras difieren para los distintos virus, y son proteínas en algunos casos (por ejemplo, picornavirus) y oligosacáridos en otros (por ejemplo, ortovirus y paramixovirus). La presencia o la ausencia de receptores desempeña una función determinante de primer orden en el tropismo celular y la patogenia viral; por ejemplo, el poliovirus es capaz de fijarse sólo en las células del sistema nervioso central y las vías digestivas de los primates. Se cree que la fijación en receptores refleja homologías configurativas fortuitas entre la estructura de la superficie del virión y un componente de la superficie celular. Por ejemplo, el virus de la inmunodeficiencia humana (HIV) se une al receptor CD4 en las células del sistema inmunitario y se cree que el virus de la rabia interactúa con los receptores de acetilcolina y que el virus de Epstein-Barr reconoce al receptor para el tercer componente del complemento en las células B. Es probable que cada célula susceptible posea por lo menos 100,000 sitios receptores para un virus dado.
Después de la fijación, la partícula viral entra en la célula. Esta etapa se conoce como penetración, viropexia o fagocitosis. En algunos sistemas, se realiza por endocitosis mediada por receptores, con captación de las partículas virales ingeridas dentro de endosomas. Están menos claros los detalles de la penetración en otros sistemas. Ocurre perdida de la cubierta de manera concomitante con la penetración o poco después de la misma. La pérdida de la cubierta, o descubrimiento, es la separación física del ácido nucleico viral (o, en algunos casos, de las nucleocápsides inteARNs) a partir de los componentes estructurales externos del virión. En este momento se pierde la infecciosidad del virus original. Los virus son los únicos agentes infecciosos en los que la disolución del agente infectante es una etapa obligatoria de la vía de la replicación. Poco después de quedar descubierto el genoma viral, se inicia la fase sintética del ciclo de replicación del virus. El aspecto esencial de la replicación viral consiste en que deben transcribirse los ARNm específicos del ácido nucleico viral para que se exprese y duplique con éxito la información genética. Una vez que se logra esta etapa, los virus recurren a los componentes de la célula para traducir el ARNm. Diversas clases de virus emplean vías diferentes para sintetizar los ARNm según la estructura del ácido nucleico viral. Algunos virus (por ejemplo, rhabdovirus, ortomixovirus y paramixovirus) poseen ARN polimerasas que sintetizan a los ARNm. Los virus ARN de este tipo se llaman virus de tira negativa (de sentido negativo), puesto que su genoma de ARN de una sola tira es complementario del ARNm, el cual se designa por acuerdo general como de tira positiva (o de sentido positivo). Durante la replicación viral se sintetizan todas las macromoléculas especificadas del virus en una secuencia muy organizada. En algunas infecciones por virus, notablemente las producidas por virus de doble tira que contienen ADN, se sintetizan proteínas virales tempranas poco después de la infección y aparecen proteínas tardías sólo durante la fase tardía del proceso infeccioso, una vez que ha ocurrido la síntesis del ADN viral. Pueden activarse o no genes tempranos cuando se elaboran los productos tardíos. En contraste, se expresa al mismo tiempo la mayor parte de la información genética de los virus que contienen ARN o toda ella. Además de estos controles temporales, existen también controles cuantitativos puesto que no se elaboran todas las proteínas virales en las mismas cantidades. Las proteínas específicas del virus pueden regular la extensión de la transcripción del genoma o la traducción del ARNm viral.
Se encuentra la variación más amplia en las estrategias de la expresión genética entre los virus que contienen ARN. Algunos viriones poseen polimerasas (ortomixovirus, reovirus); otros sistemas emplean mensajes subgenómicos generados a veces por empalme (ortomixovirus, retrovirus); por último, algunos virus sintetizan grandes precursores poliproteínicos que se someten a procesamiento y segmentación para generar los productos génicos finales (picornavirus, retrovirus). La extensión a la que las enzimas específicas del virus participan en estos procesos varía de un grupo a otro. En general, los virus de mayor tamaño (herpesvirus, poxvirus) son más independientes de las funciones celulares que los virus de menor tamaño. Esta es una razón de que los virus de mayor tamaño sean más sensibles a la quimioterapia antiviral, porque se dispone de más procesos específicos de los virus como blancos de la acción farmacológica. Los sitios intracelulares en los que ocurren los diferentes acontecimientos de la replicación viral, varían de un grupo a otro. Es posible plantear algunas generalizaciones. La proteína viral se sintetiza en el citoplasma, sobre los polirribosomas compuestos por ARNm específico del virus y ribosomas de la célula huésped. El ADN viral suele replicarse en el núcleo. El ARN genómico viral se duplica generalmente en el citoplasma celular, aunque hay excepciones.
89316 Replicación y maduración Poco después de quedar descubierto el genoma viral, se inicia la fase sintética del ciclo de replicación del virus. El aspecto esencial de la replicación viral consiste en que deben transcribirse los ARNm específicos del ácido nucleico viral para que se exprese y duplique con éxito la información genética. Una vez que se logra esta etapa, los virus recurren a los componentes de la célula para traducir el ARNm. Diversas clases de virus emplean vías diferentes para sintetizar los ARNm según la estructura del ácido nucleico viral. Algunos virus (por ejemplo, rhabdovirus, ortomixovirus y paramixovirus) poseen ARN polimerasas que sintetizan a los ARNm. Los virus ARN de este tipo se llaman virus de tira negativa (de sentido negativo), puesto que su genoma de ARN de una sola tira es complementario del ARNm, el cual se designa por acuerdo general como de tira positiva (o de sentido positivo). Durante la replicación viral se sintetizan todas las macromoléculas especificadas del virus en una secuencia muy organizada. En algunas infecciones por virus, notablemente las producidas por virus de doble tira que contienen ADN, se sintetizan proteínas virales tempranas poco después de la infección y aparecen proteínas tardías sólo durante la fase tardía del proceso infeccioso, una vez que ha ocurrido la síntesis del ADN viral. Pueden activarse o no genes tempranos cuando se elaboran los productos tardíos. En contraste, se expresa al mismo tiempo la mayor parte de la información genética de los virus que contienen ARN o toda ella. Además de estos controles temporales, existen también controles cuantitativos puesto que no se elaboran todas las proteínas virales en las mismas cantidades. Las proteínas específicas del virus pueden regular la extensión de la transcripción del genoma o la traducción del ARNm viral.
89316 Buenos modelos para el estudio de la expresión genética son los virus y los bacteriófagos pequeños de los animales. Su pequeño tamaño ha permitido aclarar la secuencia total de nucleótidos de unos cuantos virus. Esto ha hecho que se descubran genes sobrepuestos en los que se emplean algunas secuencias de ADN para la síntesis de dos poli péptidos diferentes, ya sea mediante el empleo de dos armazones de lectura diferentes o por medio de dos moléculas de ARNm que emplean los mismos armazones de lectura pero puntos de iniciación diferentes. Un sistema viral (adenovirus) reveló por primera vez el fenómeno de procesamiento de ARNm que se llama "empalme", por medio del cual se generan las secuencias de ARNm que codifican una proteína determinada a partir de secuencias separadas del modelo, con empalme de las secuencias no codificadoras intermedias de la transcripción. Se encuentra la variación más amplia en las estrategias de la expresión genética entre los virus que contienen ARN. Algunos viriones poseen polimerasas (ortomixovirus, reovirus); otros sistemas emplean mensajes subgenómicos generados a veces por empalme (ortomixovirus, retrovirus); por último, algunos virus sintetizan grandes precursores poliproteínicos que se someten a procesamiento y segmentación para generar los productos génicos finales (picornavirus, retrovirus). La extensión a la que las enzimas específicas del virus participan en estos procesos varía de un grupo a otro. En general, los virus de mayor tamaño (herpesvirus, poxvirus) son más independientes de las funciones celulares que los virus de menor tamaño. Esta es una razón de que los virus de mayor tamaño sean más sensibles a la quimioterapia antiviral, porque se dispone de más procesos específicos de los virus como blancos de la acción farmacológica.
Los sitios intracelulares en los que ocurren los diferentes acontecimientos de la replicación viral, varían de un grupo a otro. Es posible plantear algunas generalizaciones. La proteína viral se sintetiza en el citoplasma, sobre los poli ribosomas compuestos por ARNm específico del virus y ribosomas de la célula huésped. El ADN viral suele replicarse en el núcleo. El ARN genómico viral se duplica generalmente en el citoplasma celular, aunque hay excepciones.
89316 Maduración y salida de virus desnudos y envueltos En las etapas tardías del ciclo de multiplicación viral las moléculas de ácido nucleico y proteínas virales sintetizadas en la célula infectada deben ensamblarse para formar los nuevos viriones y salir al espacio extracelular para estar en condiciones de infectar nuevas células. Este proceso de maduración y ensamble, también denominado morfogénesis, comprende el armado de la cápside a partir de las unidades estructurales y luego la formación del virión completo. El mecanismo va a diferir en base a dos propiedades del virión: la estructura de la cápside y la presencia o no de envoltura. La maduración: El mecanismo mediante el cual las cápsidas se ensamblan alrededor de losácidos nucleicos no se conoce tan bien como la maduración de los fagos. Parece ser un proceso espontáneo en el que podrían estar implicadas algunas proteínas de la célula hospedadora. La liberación: Los virus envueltos adquieran su cubierta en el proceso de salida de la célula hospedadora. Antes de que el virus deje la célula, los genes víricos hacen que la célula hospedadora sintetice las proteínas quedan embebidas en la región de la membrana plasmática de la célula, que constituirá la cubierta vírica. El virus se sitúa debajo de dicha región de la membrana plasmática de la célula, que constituirá la cubierta vírica. El virus se sitúa debajo de dicha región de la membrana; la empuja hacia fuera formando una yema o protuberancia y finalmente, sale de la célula mediante un proceso de exocitosis. Como consecuencia de esta salida, el virión de un virus envuelto es una estructura compuesta; es decir, el contenido del virión y la mayor parte de las proteínas de su membrana son codificados por el genoma vírico, pero la porción lípida está codificada por el genoma de la célula hospedadora. Los virus desnudos se liberan por un mecanismo más simple, similar al que realiza el fago T4.Finalizad el ensamblaje, los viriones se acumulan en el citoplasma, la célula se isa y muere liberando los viriones.
89316 Salida de un virus envuelto Los genes víricos no se expresan hasta que el virus pierde sus cubiertas. Las membranas de algunos virus envueltos y las cápsidas de la mayoría de los virus desnudos son eliminados durante el proceso de penetración. Los viriones que penetran parcial o totalmente intactos son desnudados en el interior de la célula, por medio de enzimas hidrolíticas del hospedador, generalmente enzimas lisosómicas.
yessenia severino sanchez mat:89204 ADHESION El primer paso en la infección de una célula es la adhesión o fijación a la superficie celular. Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula. Las células que carecen de los receptores apropiados no son susceptibles al virus. PENETRACIÓN Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus.
Virus envueltos
(A) Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular.
(B) Entada vía endosomas en la superficie celular.
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
Virus no envueltos o desnudos Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras. EMSAMBLAJE Y MADURACION Ensamblaje de los componentes de los virus para formar nuevas partículas víricas (proceso también llamado maduración).
Los virus desnudos realizan su ensamblaje y maduración en forma intracelular y adquieren capacidad infectante dentro de la célula. Para su salida requieren de la desintegración celular. Los virus envueltos en el último paso al ensamblaje el virion se une con la membrana celular a la salida de la célula, mediante proteínas que se insertan en la superficie interna y exterior de la membrana. El virion naciente brota atraves de la membrana. El ensamblaje y maduración con la expulsión atraves de la superficie celular proporcionan un mecanismo más eficaz de salida, ya que no depende de la desintegración de la célula infectada. SALIDA Liberación de los nuevos virus de la célula hospedadora, que ya pueden infectar otras células. Pueden producirse con o sin rotura de la célula, dependiendo de los virus. Los virus desnudos salen por lisis celular lo contrario de los envueltos que salen por gemación.
Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada
La única característica de la multiplicación viral consiste en que, poco después de su interacción con una célula huésped, el virión infectante se desintegra y pierde la infecciosidad que se puede medir. Esta fase del ciclo celular se llama periodo de eclipse; su duración varía dependiendo del virus y de la célula huésped en particular, y termina con la formación de las primeras partículas virales descendientes infecciosas. El periodo de eclipse es, en realidad, de intensa actividad sintética, puesto que la célula se redirige hacia la satisfacción de las necesidades del "pirata" viral. En algunos casos, tan pronto como el ácido nucleico viral penetra a la célula huésped, el metabolismo celular es recanalizado exclusivamente hacia la síntesis de nuevas partículas virales. En otros casos, los procesos metabólicos de la célula huésped no se alteran significativamente, aunque la célula sintetiza proteínas virales y ácidos nucleicos. Los virus han desarrollado diversas estrategias para lograr la multiplicación en las células huéspedes parasitadas por ellos. Aunque los detalles varían de un grupo a otro, el patrón general de los ciclos de replicación es semejante. Virus no envueltos o desnudos Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras. -La primera etapa de la infección viral es la interacción de un virión con un sitio receptor específica de la superficie celular. Las moléculas receptoras difieren para los distintos virus, y son proteínas en algunos casos (por ejemplo, picornavirus) y oligosacáridos en otros (por ejemplo, ortovirus y paramixovirus). La presencia o la ausencia de receptores desempeña una función determinante de primer orden en el tropismo celular y la patogenia viral; por ejemplo, el poliovirus es capaz de fijarse sólo en las células del sistema nervioso central y las vías digestivas de los primates. Se cree que la fijación en receptores refleja homologías configurativas fortuitas entre la estructura de la superficie del virión y un componente de la superficie celular. Por ejemplo, el virus de la inmunodeficiencia humana (HIV) se une al receptor CD4 en las células del sistema inmunitario y se cree que el virus de la rabia interactúa con los receptores de acetilcolina y que el virus de Epstein-Barr reconoce al receptor para el tercer componente del complemento en las células B. Es probable que cada célula susceptible posea por lo menos 100,000 sitios receptores para un virus dado
-mediante Después de la fijación, la partícula viral entra en la célula. Esta etapa se conoce como penetración, viropexia o fagocitosis. En algunos sistemas, se realiza por endocitosis mediada por receptores, con captación de las partículas virales ingeridas dentro de endosomas. Están menos claros los detalles de la penetración en otros sistemas. Ocurre perdida de la cubierta de manera concomitante con la penetración o poco después de la misma. La pérdida de la cubierta, o descubrimiento, es la separación física del ácido nucleico viral (o, en algunos casos, de las nucleocápsides inteARNs) a partir de los componentes estructurales externos del virión. En este momento se pierde la infecciosidad del virus original. Los virus son los únicos agentes infecciosos en los que la disolución del agente infectante es una etapa obligatoria de la vía de la replicación



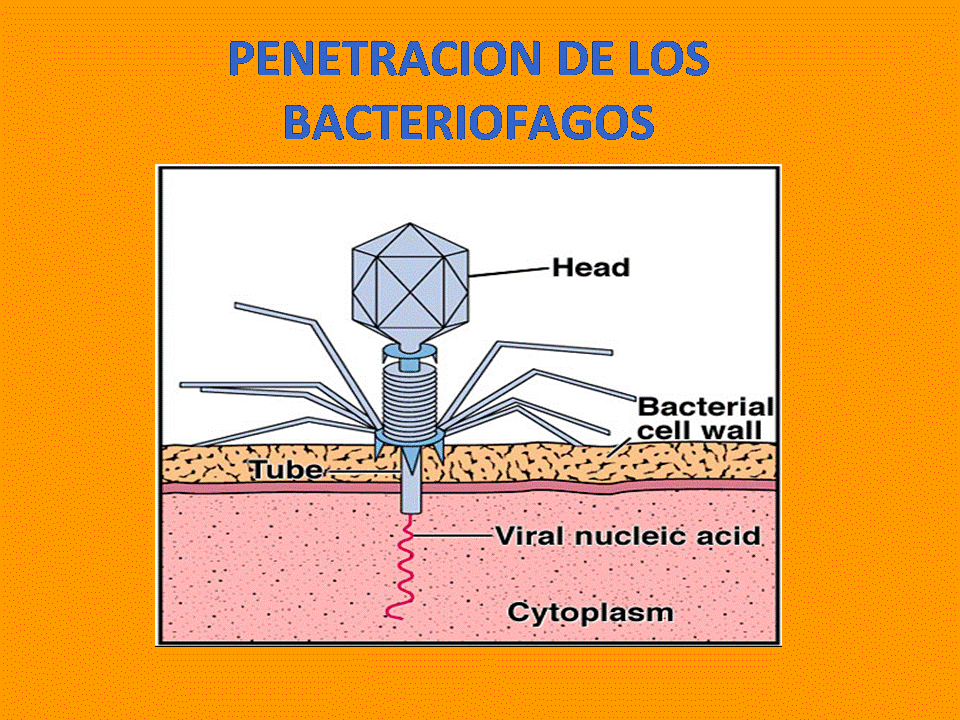



Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada.
ResponderEliminarLa única característica de la multiplicación viral consiste en que, poco después de su interacción con una célula huésped, el virión infectante se desintegra y pierde la infecciosidad que se puede medir. Esta fase del ciclo celular se llama periodo de eclipse; su duración varía dependiendo del virus y de la célula huésped en particular, y termina con la formación de las primeras partículas virales descendientes infecciosas. El periodo de eclipse es, en realidad, de intensa actividad sintética, puesto que la célula se redirige hacia la satisfacción de las necesidades del "pirata" viral. En algunos casos, tan pronto como el ácido nucleico viral penetra a la célula huésped, el metabolismo celular es recanalizado exclusivamente hacia la síntesis de nuevas partículas virales. En otros casos, los procesos metabólicos de la célula huésped no se alteran significativamente, aunque la célula sintetiza proteínas virales y ácidos nucleicos.
ResponderEliminarLos virus han desarrollado diversas estrategias para lograr la multiplicación en las células huéspedes parasitadas por ellos. Aunque los detalles varían de un grupo a otro, el patrón general de los ciclos de replicación es semejante.
ResponderEliminarLa primera etapa de la infección viral es la interacción de un virión con un sitio receptor específica de la superficie celular. Las moléculas receptoras difieren para los distintos virus, y son proteínas en algunos casos (por ejemplo, picornavirus) y oligosacáridos en otros (por ejemplo, ortovirus y paramixovirus). La presencia o la ausencia de receptores desempeña una función determinante de primer orden en el tropismo celular y la patogenia viral; por ejemplo, el poliovirus es capaz de fijarse sólo en las células del sistema nervioso central y las vías digestivas de los primates. Se cree que la fijación en receptores refleja homologías configurativas fortuitas entre la estructura de la superficie del virión y un componente de la superficie celular. Por ejemplo, el virus de la inmunodeficiencia humana (HIV) se une al receptor CD4 en las células del sistema inmunitario y se cree que el virus de la rabia interactúa con los receptores de acetilcolina y que el virus de Epstein-Barr reconoce al receptor para el tercer componente del complemento en las células B. Es probable que cada célula susceptible posea por lo menos 100,000 sitios receptores para un virus dado.
ResponderEliminarDespués de la fijación, la partícula viral entra en la célula. Esta etapa se conoce como penetración, viropexia o fagocitosis. En algunos sistemas, se realiza por endocitosis mediada por receptores, con captación de las partículas virales ingeridas dentro de endosomas. Están menos claros los detalles de la penetración en otros sistemas. Ocurre perdida de la cubierta de manera concomitante con la penetración o poco después de la misma. La pérdida de la cubierta, o descubrimiento, es la separación física del ácido nucleico viral (o, en algunos casos, de las nucleocápsides inteARNs) a partir de los componentes estructurales externos del virión. En este momento se pierde la infecciosidad del virus original.
ResponderEliminarLos virus son los únicos agentes infecciosos en los que la disolución del agente infectante es una etapa obligatoria de la vía de la replicación.
Poco después de quedar descubierto el genoma viral, se inicia la fase sintética del ciclo de replicación del virus. El aspecto esencial de la replicación viral consiste en que deben transcribirse los ARNm específicos del ácido nucleico viral para que se exprese y duplique con éxito la información genética. Una vez que se logra esta etapa, los virus recurren a los componentes de la célula para traducir el ARNm. Diversas clases de virus emplean vías diferentes para sintetizar los ARNm según la estructura del ácido nucleico viral. Algunos virus (por ejemplo, rhabdovirus, ortomixovirus y paramixovirus) poseen ARN polimerasas que sintetizan a los ARNm. Los virus ARN de este tipo se llaman virus de tira negativa (de sentido negativo), puesto que su genoma de ARN de una sola tira es complementario del ARNm, el cual se designa por acuerdo general como de tira positiva (o de sentido positivo).
ResponderEliminarDurante la replicación viral se sintetizan todas las macromoléculas especificadas del virus en una secuencia muy organizada. En algunas infecciones por virus, notablemente las producidas por virus de doble tira que contienen ADN, se sintetizan proteínas virales tempranas poco después de la infección y aparecen proteínas tardías sólo durante la fase tardía del proceso infeccioso, una vez que ha ocurrido la síntesis del ADN viral. Pueden activarse o no genes tempranos cuando se elaboran los productos tardíos. En contraste, se expresa al mismo tiempo la mayor parte de la información genética de los virus que contienen ARN o toda ella. Además de estos controles temporales, existen también controles cuantitativos puesto que no se elaboran todas las proteínas virales en las mismas cantidades. Las proteínas específicas del virus pueden regular la extensión de la transcripción del genoma o la traducción del ARNm viral.
Se encuentra la variación más amplia en las estrategias de la expresión genética entre los virus que contienen ARN. Algunos viriones poseen polimerasas (ortomixovirus, reovirus); otros sistemas emplean mensajes subgenómicos generados a veces por empalme (ortomixovirus, retrovirus); por último, algunos virus sintetizan grandes precursores poliproteínicos que se someten a procesamiento y segmentación para generar los productos génicos finales (picornavirus, retrovirus).
ResponderEliminarLa extensión a la que las enzimas específicas del virus participan en estos procesos varía de un grupo a otro. En general, los virus de mayor tamaño (herpesvirus, poxvirus) son más independientes de las funciones celulares que los virus de menor tamaño. Esta es una razón de que los virus de mayor tamaño sean más sensibles a la quimioterapia antiviral, porque se dispone de más procesos específicos de los virus como blancos de la acción farmacológica.
Los sitios intracelulares en los que ocurren los diferentes acontecimientos de la replicación viral, varían de un grupo a otro. Es posible plantear algunas generalizaciones. La proteína viral se sintetiza en el citoplasma, sobre los polirribosomas compuestos por ARNm específico del virus y ribosomas de la célula huésped. El ADN viral suele replicarse en el núcleo. El ARN genómico viral se duplica generalmente en el citoplasma celular, aunque hay excepciones.
Diapositiva #2
ResponderEliminar1. ADHESION
El primer paso en la infección de una célula es la adhesión o fijación a la superficie celular. Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula. Las células que carecen de los receptores apropiados no son susceptibles al virus.
Diapositiva #3
ResponderEliminar2. PENETRACIÓN
Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus.
Virus envueltos
(A) Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular.
(B) Entada vía endosomas en la superficie celular.
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
Virus no envueltos o desnudos
Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
Diapositiva #5
ResponderEliminar3. EMSAMBLAJE Y MADURACION
Ensamblaje de los componentes de los virus para formar nuevas partículas víricas (proceso también llamado maduración).
Los virus desnudos realizan su ensamblaje y maduración en forma intracelular y adquieren capacidad infectante dentro de la célula. Para su salida requieren de la desintegración celular. Los virus envueltos en el último paso al ensamblaje el virion se une con la membrana celular a la salida de la célula, mediante proteínas que se insertan en la superficie interna y exterior de la membrana. El virion naciente brota atraves de la membrana. El ensamblaje y maduración con la expulsión atraves de la superficie celular proporcionan un mecanismo más eficaz de salida, ya que no depende de la desintegración de la célula infectada.
Diapositiva #7
ResponderEliminar4. SALIDA
Liberación de los nuevos virus de la célula hospedadora, que ya pueden infectar otras células. Pueden producirse con o sin rotura de la célula, dependiendo de los virus. Los virus desnudos salen por lisis celular lo contrario de los envueltos que salen por gemación.
paula ortiz 88402
ResponderEliminar1. Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada
88402
ResponderEliminarLa única característica de la multiplicación viral consiste en que, poco después de su interacción con una célula huésped, el virión infectante se desintegra y pierde la infecciosidad que se puede medir. Esta fase del ciclo celular se llama periodo de eclipse; su duración varía dependiendo del virus y de la célula huésped en particular, y termina con la formación de las primeras partículas virales descendientes infecciosas. El periodo de eclipse es, en realidad, de intensa actividad sintética, puesto que la célula se redirige hacia la satisfacción de las necesidades del "pirata" viral. En algunos casos, tan pronto como el ácido nucleico viral penetra a la célula huésped, el metabolismo celular es recanalizado exclusivamente hacia la síntesis de nuevas partículas virales. En otros casos, los procesos metabólicos de la célula huésped no se alteran significativamente, aunque la célula sintetiza proteínas virales y ácidos nucleicos.
88402
ResponderEliminarLos virus han desarrollado diversas estrategias para lograr la multiplicación en las células huéspedes parasitadas por ellos. Aunque los detalles varían de un grupo a otro, el patrón general de los ciclos de replicación es semejante.
Virus no envueltos o desnudos
Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
88402
ResponderEliminar4-La primera etapa de la infección viral es la interacción de un virión con un sitio receptor específica de la superficie celular. Las moléculas receptoras difieren para los distintos virus, y son proteínas en algunos casos (por ejemplo, picornavirus) y oligosacáridos en otros (por ejemplo, ortovirus y paramixovirus). La presencia o la ausencia de receptores desempeña una función determinante de primer orden en el tropismo celular y la patogenia viral; por ejemplo, el poliovirus es capaz de fijarse sólo en las células del sistema nervioso central y las vías digestivas de los primates. Se cree que la fijación en receptores refleja homologías configurativas fortuitas entre la estructura de la superficie del virión y un componente de la superficie celular. Por ejemplo, el virus de la inmunodeficiencia humana (HIV) se une al receptor CD4 en las células del sistema inmunitario y se cree que el virus de la rabia interactúa con los receptores de acetilcolina y que el virus de Epstein-Barr reconoce al receptor para el tercer componente del complemento en las células B. Es probable que cada célula susceptible posea por lo menos 100,000 sitios receptores para un virus dado
88402
ResponderEliminar5-mediante Después de la fijación, la partícula viral entra en la célula. Esta etapa se conoce como penetración, viropexia o fagocitosis. En algunos sistemas, se realiza por endocitosis mediada por receptores, con captación de las partículas virales ingeridas dentro de endosomas. Están menos claros los detalles de la penetración en otros sistemas. Ocurre perdida de la cubierta de manera concomitante con la penetración o poco después de la misma. La pérdida de la cubierta, o descubrimiento, es la separación física del ácido nucleico viral (o, en algunos casos, de las nucleocápsides inteARNs) a partir de los componentes estructurales externos del virión. En este momento se pierde la infecciosidad del virus original.
Los virus son los únicos agentes infecciosos en los que la disolución del agente infectante es una etapa obligatoria de la vía de la replicación
88402
ResponderEliminar6-durante Poco después de quedar descubierto el genoma viral, se inicia la fase sintética del ciclo de replicación del virus. El aspecto esencial de la replicación viral consiste en que deben transcribirse los ARNm específicos del ácido nucleico viral para que se exprese y duplique con éxito la información genética. Una vez que se logra esta etapa, los virus recurren a los componentes de la célula para traducir el ARNm. Diversas clases de virus emplean vías diferentes para sintetizar los ARNm según la estructura del ácido nucleico viral. Algunos virus (por ejemplo, rhabdovirus, ortomixovirus y paramixovirus) poseen ARN polimerasas que sintetizan a los ARNm. Los virus ARN de este tipo se llaman virus de tira negativa (de sentido negativo), puesto que su genoma de ARN de una sola tira es complementario del ARNm, el cual se designa por acuerdo general como de tira positiva (o de sentido positivo).
Durante la replicación viral se sintetizan todas las macromoléculas especificadas del virus en una secuencia muy organizada. En algunas infecciones por virus, notablemente las producidas por virus de doble tira que contienen ADN, se sintetizan proteínas virales tempranas poco después de la infección y aparecen proteínas tardías sólo durante la fase tardía del proceso infeccioso, una vez que ha ocurrido la síntesis del ADN viral. Pueden activarse o no genes tempranos cuando se elaboran los productos tardíos. En contraste, se expresa al mismo tiempo la mayor parte de la información genética de los virus que contienen ARN o toda ella. Además de estos controles temporales, existen también controles cuantitativos puesto que no se elaboran todas las proteínas virales en las mismas cantidades. Las proteínas específicas del virus pueden regular la extensión de la transcripción del genoma o la traducción del ARNm viral.
88402
ResponderEliminar7-todo Se encuentra la variación más amplia en las estrategias de la expresión genética entre los virus que contienen ARN. Algunos viriones poseen polimerasas (ortomixovirus, reovirus); otros sistemas emplean mensajes subgenómicos generados a veces por empalme (ortomixovirus, retrovirus); por último, algunos virus sintetizan grandes precursores poliproteínicos que se someten a procesamiento y segmentación para generar los productos génicos finales (picornavirus, retrovirus).
La extensión a la que las enzimas específicas del virus participan en estos procesos varía de un grupo a otro. En general, los virus de mayor tamaño (herpesvirus, poxvirus) son más independientes de las funciones celulares que los virus de menor tamaño. Esta es una razón de que los virus de mayor tamaño sean más sensibles a la quimioterapia antiviral, porque se dispone de más procesos específicos de los virus como blancos de la acción farmacológica.
Los sitios intracelulares en los que ocurren los diferentes acontecimientos de la replicación viral, varían de un grupo a otro. Es posible plantear algunas generalizaciones. La proteína viral se sintetiza en el citoplasma, sobre los polirribosomas compuestos por ARNm específico del virus y ribosomas de la célula huésped. El ADN viral suele replicarse en el núcleo. El ARN genómico viral se duplica generalmente en el citoplasma celular, aunque hay excepciones
Nombre: Zaida Acta. Matrícula: 86624
ResponderEliminarETAPAS DE LA REPLICACIÓN VIRAL
1- El primer paso en la infección viral es la absorción a la membrana de la célula susceptible por medio de la adhesión de ligandos virales (proteínas de la capside o glucoproteínas de las espículas, por ejemplo a receptores superficiales de la célula). La distribución en el cuerpo del hospedador de ciertos tipos de receptores celulares explica el tropismo tisular y de hospedador de los virus. Los tejidos y células que carecen de receptores específicos de virus determinados no son infectados por dichos viriones. La naturaleza de los receptores es variable y ciertos virus pueden tener más de un tipo de receptor.
2- Los virus complejos producen una rotura en la membrana celular del hospedador en uno de los puntos de anclaje, gracias a la presencia de algunas moléculas de enzimas hidrolíticas entre lasproteínas de la cápsida. A través de la rotura, el tubo central inyecta el ADN vírico, quedando la cápside vacía en el exterior de la célula diana y el ácido nucleico libre en el citoplasma. La presencia de cápsidas en la superficie celular es un buen indicio de que ha sufrido una infección vírica.
Otros virus sin envoltura lipídica se introducen en la célula con cápsida y todo, lo cual puede realizarse de dos maneras:
• Por penetración directa: después de la fijación, el virus abre una brecha en la membrana y se introduce en el citoplasma.
• Por endocitosis: la membrana forma una invaginación en torno al virus, llegando a formar una vesícula que penetra en la célula. Formada la vesícula, el virus abre una brecha en la membrana de la misma con ayuda de algunas enzimas hidrolíticas que él mismo transporta, penetrando así en el citoplasma.
Los virus con envoltura lipídica burlan la barrera de la membrana celular porque su cubierta lipídica se funde con la membrana, ya que son de la misma naturaleza. Esta fusión de membranas puede realizarse en dos lugares distintos:
• Fusión en la superficie celular: de manera que el virión penetra directamente en el citoplasma.
• Fusión con un lisosoma: se forma una vesícula por endocitosis, a la que se une un lisosoma para digerir la partícula introducida; entonces, la cubierta lipídica del virus se funde con la membrana del lisosoma y el virión escapa hacia el citoplasma.
3- Según la duración de la fase de eclipse, se suelen distinguir dos modalidades de ciclo infeccioso de un virus:
• Ciclo ordinario: el ácido nucleico vírico procede inmediatamente a la transcripción de su mensaje genético en los ARN necesarios para su multiplicación, y prosigue rápidamente el ciclo vital. Este tipo de ciclo es el más extendido en la naturaleza.
• Ciclo lisogénico: fue descubierto por André Lwoff en bacteriófagos. El ADN vírico se cierra por sus extremos generando un ADN circular. Este ADN se inserta en el ADN bacteriano en un lugar específico en el que la secuencia de nucleótidos bacterianos es semejante alguna región del ADN vírico.
Algunos virus que infectan células animales siguen también el ciclo lisogénico, como los papilomavirus de las verrugas y algunos retrovirus que producen algunos tipos de cáncer.
En el caso de los retrovirus, conviene recordar que el ácido nucleico es ARN monocatenario, por lo que la transcriptasa inversa ha de copiar el genoma vírico en forma de ADN antes de que pueda insertarse en el ADN celular.
4- La multiplicación del virus consta de la replicación de su material genético, de la transcripción de su mensaje en una molécula de ARN y de la traducción del mensaje para producir proteínasvíricas, tanto las que formarán parte de la cápsida como las proteínas enzimáticas necesarias para el ensamblaje de las piezas del virión y para algunas de las funciones anteriores. Losribosomas y la mayor parte de las enzimas que los ácidos nucleicos víricos utilizan en estos procesos son los de la célula infectada.
Este comentario ha sido eliminado por un administrador del blog.
ResponderEliminarEste comentario ha sido eliminado por un administrador del blog.
ResponderEliminarEste comentario ha sido eliminado por un administrador del blog.
ResponderEliminarEste comentario ha sido eliminado por un administrador del blog.
ResponderEliminarEste comentario ha sido eliminado por un administrador del blog.
ResponderEliminarEste comentario ha sido eliminado por un administrador del blog.
ResponderEliminarEste comentario ha sido eliminado por un administrador del blog.
ResponderEliminarEste comentario ha sido eliminado por un administrador del blog.
ResponderEliminarEste comentario ha sido eliminado por un administrador del blog.
ResponderEliminarLos virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada.
ResponderEliminarLa única característica de la multiplicación viral consiste en que, poco después de su interacción con una célula huésped, el virión infectante se desintegra y pierde la infecciosidad que se puede medir. Esta fase del ciclo celular se llama periodo de eclipse; su duración varía dependiendo del virus y de la célula huésped en particular, y termina con la formación de las primeras partículas virales descendientes infecciosas. El periodo de eclipse es, en realidad, de intensa actividad sintética, puesto que la célula se redirige hacia la satisfacción de las necesidades del "pirata" viral. En algunos casos, tan pronto como el ácido nucleico viral penetra a la célula huésped, el metabolismo celular es recanalizado exclusivamente hacia la síntesis de nuevas partículas virales. En otros casos, los procesos metabólicos de la célula huésped no se alteran significativamente, aunque la célula sintetiza proteínas virales y ácidos nucleicos.
ResponderEliminarLos virus han desarrollado diversas estrategias para lograr la multiplicación en las células huéspedes parasitadas por ellos. Aunque los detalles varían de un grupo a otro, el patrón general de los ciclos de replicación es semejante.
ResponderEliminarLa primera etapa de la infección viral es la interacción de un virión con un sitio receptor específica de la superficie celular. Las moléculas receptoras difieren para los distintos virus, y son proteínas en algunos casos (por ejemplo, picornavirus) y oligosacáridos en otros (por ejemplo, ortovirus y paramixovirus). La presencia o la ausencia de receptores desempeña una función determinante de primer orden en el tropismo celular y la patogenia viral; por ejemplo, el poliovirus es capaz de fijarse sólo en las células del sistema nervioso central y las vías digestivas de los primates. Se cree que la fijación en receptores refleja homologías configurativas fortuitas entre la estructura de la superficie del virión y un componente de la superficie celular. Por ejemplo, el virus de la inmunodeficiencia humana (HIV) se une al receptor CD4 en las células del sistema inmunitario y se cree que el virus de la rabia interactúa con los receptores de acetilcolina y que el virus de Epstein-Barr reconoce al receptor para el tercer componente del complemento en las células B. Es probable que cada célula susceptible posea por lo menos 100,000 sitios receptores para un virus dado.
Después de la fijación, la partícula viral entra en la célula. Esta etapa se conoce como penetración, viropexia o fagocitosis. En algunos sistemas, se realiza por endocitosis mediada por receptores, con captación de las partículas virales ingeridas dentro de endosomas. Están menos claros los detalles de la penetración en otros sistemas. Ocurre perdida de la cubierta de manera concomitante con la penetración o poco después de la misma. La pérdida de la cubierta, o descubrimiento, es la separación física del ácido nucleico viral (o, en algunos casos, de las nucleocápsides inteARNs) a partir de los componentes estructurales externos del virión. En este momento se pierde la infecciosidad del virus original.
ResponderEliminarLos virus son los únicos agentes infecciosos en los que la disolución del agente infectante es una etapa obligatoria de la vía de la replicación.
Poco después de quedar descubierto el genoma viral, se inicia la fase sintética del ciclo de replicación del virus. El aspecto esencial de la replicación viral consiste en que deben transcribirse los ARNm específicos del ácido nucleico viral para que se exprese y duplique con éxito la información genética. Una vez que se logra esta etapa, los virus recurren a los componentes de la célula para traducir el ARNm. Diversas clases de virus emplean vías diferentes para sintetizar los ARNm según la estructura del ácido nucleico viral. Algunos virus (por ejemplo, rhabdovirus, ortomixovirus y paramixovirus) poseen ARN polimerasas que sintetizan a los ARNm. Los virus ARN de este tipo se llaman virus de tira negativa (de sentido negativo), puesto que su genoma de ARN de una sola tira es complementario del ARNm, el cual se designa por acuerdo general como de tira positiva (o de sentido positivo).
Durante la replicación viral se sintetizan todas las macromoléculas especificadas del virus en una secuencia muy organizada. En algunas infecciones por virus, notablemente las producidas por virus de doble tira que contienen ADN, se sintetizan proteínas virales tempranas poco después de la infección y aparecen proteínas tardías sólo durante la fase tardía del proceso infeccioso, una vez que ha ocurrido la síntesis del ADN viral. Pueden activarse o no genes tempranos cuando se elaboran los productos tardíos. En contraste, se expresa al mismo tiempo la mayor parte de la información genética de los virus que contienen ARN o toda ella. Además de estos controles temporales, existen también controles cuantitativos puesto que no se elaboran todas las proteínas virales en las mismas cantidades. Las proteínas específicas del virus pueden regular la extensión de la transcripción del genoma o la traducción del ARNm viral.
Se encuentra la variación más amplia en las estrategias de la expresión genética entre los virus que contienen ARN. Algunos viriones poseen polimerasas (ortomixovirus, reovirus); otros sistemas emplean mensajes subgenómicos generados a veces por empalme (ortomixovirus, retrovirus); por último, algunos virus sintetizan grandes precursores poliproteínicos que se someten a procesamiento y segmentación para generar los productos génicos finales (picornavirus, retrovirus).
ResponderEliminarLa extensión a la que las enzimas específicas del virus participan en estos procesos varía de un grupo a otro. En general, los virus de mayor tamaño (herpesvirus, poxvirus) son más independientes de las funciones celulares que los virus de menor tamaño. Esta es una razón de que los virus de mayor tamaño sean más sensibles a la quimioterapia antiviral, porque se dispone de más procesos específicos de los virus como blancos de la acción farmacológica.
Los sitios intracelulares en los que ocurren los diferentes acontecimientos de la replicación viral, varían de un grupo a otro. Es posible plantear algunas generalizaciones. La proteína viral se sintetiza en el citoplasma, sobre los polirribosomas compuestos por ARNm específico del virus y ribosomas de la célula huésped. El ADN viral suele replicarse en el núcleo. El ARN genómico viral se duplica generalmente en el citoplasma celular, aunque hay excepciones.
Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada.
ResponderEliminarLa única característica de la multiplicación viral consiste en que, poco después de su interacción con una célula huésped, el virión infectante se desintegra y pierde la infecciosidad que se puede medir. Esta fase del ciclo celular se llama periodo de eclipse; su duración varía dependiendo del virus y de la célula huésped en particular, y termina con la formación de las primeras partículas virales descendientes infecciosas. El periodo de eclipse es, en realidad, de intensa actividad sintética, puesto que la célula se redirige hacia la satisfacción de las necesidades del "pirata" viral. En algunos casos, tan pronto como el ácido nucleico viral penetra a la célula huésped, el metabolismo celular es recanalizado exclusivamente hacia la síntesis de nuevas partículas virales. En otros casos, los procesos metabólicos de la célula huésped no se alteran significativamente, aunque la célula sintetiza proteínas virales y ácidos nucleicos.
mat:89206
Los virus han desarrollado diversas estrategias para lograr la multiplicación en las células huéspedes parasitadas por ellos. Aunque los detalles varían de un grupo a otro, el patrón general de los ciclos de replicación es semejante
ResponderEliminarLa primera etapa de la infección viral es la interacción de un virión con un sitio receptor específica de la superficie celular. Las moléculas receptoras difieren para los distintos virus, y son proteínas en algunos casos (por ejemplo, picornavirus) y oligosacáridos en otros (por ejemplo, ortovirus y paramixovirus). La presencia o la ausencia de receptores desempeña una función determinante de primer orden en el tropismo celular y la patogenia viral; por ejemplo, el poliovirus es capaz de fijarse sólo en las células del sistema nervioso central y las vías digestivas de los primates. Se cree que la fijación en receptores refleja homologías configurativas fortuitas entre la estructura de la superficie del virión y un componente de la superficie celular. Por ejemplo, el virus de la inmunodeficiencia humana (HIV) se une al receptor CD4 en las células del sistema inmunitario y se cree que el virus de la rabia interactúa con los receptores de acetilcolina y que el virus de Epstein-Barr reconoce al receptor para el tercer componente del complemento en las células B. Es probable que cada célula susceptible posea por lo menos 100,000 sitios receptores para un virus dado.
mat:89206
Después de la fijación, la partícula viral entra en la célula. Esta etapa se conoce como penetración, viropexia o fagocitosis. En algunos sistemas, se realiza por endocitosis mediada por receptores, con captación de las partículas virales ingeridas dentro de endosomas. Están menos claros los detalles de la penetración en otros sistemas. Ocurre perdida de la cubierta de manera concomitante con la penetración o poco después de la misma. La pérdida de la cubierta, o descubrimiento, es la separación física del ácido nucleico viral (o, en algunos casos, de las nucleocápsides inteARNs) a partir de los componentes estructurales externos del virión. En este momento se pierde la infecciosidad del virus original.
ResponderEliminarLos virus son los únicos agentes infecciosos en los que la disolución del agente infectante es una etapa obligatoria de la vía de la replicación.
Poco después de quedar descubierto el genoma viral, se inicia la fase sintética del ciclo de replicación del virus. El aspecto esencial de la replicación viral consiste en que deben transcribirse los ARNm específicos del ácido nucleico viral para que se exprese y duplique con éxito la información genética. Una vez que se logra esta etapa, los virus recurren a los componentes de la célula para traducir el ARNm. Diversas clases de virus emplean vías diferentes para sintetizar los ARNm según la estructura del ácido nucleico viral. Algunos virus (por ejemplo, rhabdovirus, ortomixovirus y paramixovirus) poseen ARN polimerasas que sintetizan a los ARNm. Los virus ARN de este tipo se llaman virus de tira negativa (de sentido negativo), puesto que su genoma de ARN de una sola tira es complementario del ARNm, el cual se designa por acuerdo general como de tira positiva (o de sentido positivo).
Durante la replicación viral se sintetizan todas las macromoléculas especificadas del virus en una secuencia muy organizada. En algunas infecciones por virus, notablemente las producidas por virus de doble tira que contienen ADN, se sintetizan proteínas virales tempranas poco después de la infección y aparecen proteínas tardías sólo durante la fase tardía del proceso infeccioso, una vez que ha ocurrido la síntesis del ADN viral. Pueden activarse o no genes tempranos cuando se elaboran los productos tardíos. En contraste, se expresa al mismo tiempo la mayor parte de la información genética de los virus que contienen ARN o toda ella. Además de estos controles temporales, existen también controles cuantitativos puesto que no se elaboran todas las proteínas virales en las mismas cantidades. Las proteínas específicas del virus pueden regular la extensión de la transcripción del genoma o la traducción del ARNm viral.
mat:89206
Se encuentra la variación más amplia en las estrategias de la expresión genética entre los virus que contienen ARN. Algunos viriones poseen polimerasas (ortomixovirus, reovirus); otros sistemas emplean mensajes subgenómicos generados a veces por empalme (ortomixovirus, retrovirus); por último, algunos virus sintetizan grandes precursores poliproteínicos que se someten a procesamiento y segmentación para generar los productos génicos finales (picornavirus, retrovirus).
ResponderEliminarLa extensión a la que las enzimas específicas del virus participan en estos procesos varía de un grupo a otro. En general, los virus de mayor tamaño (herpesvirus, poxvirus) son más independientes de las funciones celulares que los virus de menor tamaño. Esta es una razón de que los virus de mayor tamaño sean más sensibles a la quimioterapia antiviral, porque se dispone de más procesos específicos de los virus como blancos de la acción farmacológica.
Los sitios intracelulares en los que ocurren los diferentes acontecimientos de la replicación viral, varían de un grupo a otro. Es posible plantear algunas generalizaciones. La proteína viral se sintetiza en el citoplasma, sobre los polirribosomas compuestos por ARNm específico del virus y ribosomas de la célula huésped. El ADN viral suele replicarse en el núcleo. El ARN genómico viral se duplica generalmente en el citoplasma celular, aunque hay excepciones.
mat:89206
Diapositiba1
ResponderEliminarEtapas de la replicación viral
La replicación es el proceso en el cual se copia el ADN. Este proceso es semiconservativo y bidireccional. Funciona igual, tanto en procariontes como en eucarionte, salvo algunos cambio, principalmente de las proteínas que participan. Se dieron muchas hipótesis sobre como se dupllicaba el ADN hasta que Watson y Crick propusieron la hipótesis semiconservativa, según la cual, las nuevas moléculas de ADN formadas a partir de otra antigua, tienen una hebra antigua y otra nueva. La replicación ocurre en 3 etapas: La iniciación La elongación La terminación.
Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada.
El ciclo de replicación de los virus es el término utilizado para describir el ciclo de reproducción de los virus. Este consta generalmente de las siguientes fases: fijación y entrada en la célula, eclipse, multiplicación y liberación del virus. Los virus son formas acelulares que no pueden reproducirse por si mismos en el exterior; para ello requieren introducirse en células donde controlarán sus mecanismos reproductivos.
Diapositiva2
ResponderEliminarADHESION
El primer paso en la infección de una célula es la adhesión o fijación a la superficie celular. Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula. Las células que carecen de los receptores apropiados no son susceptibles al virus.
Diapositiva3
ResponderEliminarPENETRACIÓN
Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus.
Virus envueltos
(A) Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular.
(B) Entada vía endosomas en la superficie celular.
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
Virus no envueltos o desnudos
Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
Diapositiva4
ResponderEliminar3. EMSAMBLAJE Y MADURACION
Ensamblaje de los componentes de los virus para formar nuevas partículas víricas (proceso también llamado maduración).
Los virus desnudos realizan su ensamblaje y maduración en forma intracelular y adquieren capacidad infectante dentro de la célula. Para su salida requieren de la desintegración celular. Los virus envueltos en el último paso al ensamblaje el virion se une con la membrana celular a la salida de la célula, mediante proteínas que se insertan en la superficie interna y exterior de la membrana. El virion naciente brota atraves de la membrana. El ensamblaje y maduración con la expulsión atraves de la superficie celular proporcionan un mecanismo más eficaz de salida, ya que no depende de la desintegración de la célula infectada.
Diapositiva5
ResponderEliminarSALIDA
Liberación de los nuevos virus de la célula hospedadora, que ya pueden infectar otras células. Pueden producirse con o sin rotura de la célula, dependiendo de los virus. Los virus desnudos salen por lisis celular lo contrario de los envueltos que salen por gemación. La distribución en el cuerpo del hospedador de ciertos tipos de receptores celulares explica el tropismo tisular y de hospedador de los virus. Los tejidos y células que carecen de receptores específicos de virus determinados no son infectados por dichos viriones. La naturaleza de los receptores es variable y ciertos virus pueden tener más de un tipo de receptor.
Matricula 75596
ResponderEliminarEtapas de la replicacion viral:
EVENTOS PRINCIPALES EN LA REPLICACIÓN
1. ADSORPCIÓN
El primer paso en la infección de una célula es la adhesión o fijación a la superficie celular. Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula. Las células que carecen de los receptores apropiados no son susceptibles al virus.
2. PENETRACIÓN
Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus.
Virus envueltos
(A) Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular. (Figura 1).
(B) Entada vía endosomas en la superficie celular (figura 2)
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
Virus no envueltos o desnudos
Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
3. PÉRDIDA DE LA CÁPSULA
El ácido nucleido debe de estar lo suficientemente expuesto como para que la replicación viral comience en esta fase. Cuando el ácido nucleico es expuesto, las partículas víricas infecciosas no pueden removerse de las células – este es el comienz de la fase de ECLIPSE – la cual perdura hasta que viriones infecciosos nuevos sean creados.
4. SÍNTESIS DE ÁCIDO NUCLEICO Y PROTEINAS VIRALES
Existen diversos mecanismos, algunos seran abarcados en capítulos posteriores.
5. ENSAMBLAJE/MADURACIÓN
Nuevas partículas víricas son ensambladas. Puede haber un estadio de maduración posterior al proceso inicial de ensamblaje.
6. LIBERACIÓN O DESCARGA
Los virus pueden ser liberados mediante lisis celular, o, si son envueltos, pueden yemar de las células. Los virus que yeman (figuras 3 y 4) no necesariamente destruyen la célula. Por tanto, algunos de estos virus son capaces de instaurar infecciones persistentes. No todas las partículas víricas liberadas son infecciosas. La proporción partículas infecciosas:partículas no-infecciosas varía según el virus y las condiciones de crecimiento.
arlin aracelis silvestre s 89308
ResponderEliminarDiaposiiva 1
ETAPAS DE LA REPLICACIÓN VIRAL
El primer paso en la infección viral es la absorción a la membrana de la célula susceptible por medio de la adhesión de ligandos virales (proteínas de la capside o glucoproteínas de las espículas, por ejemplo a receptores superficiales de la célula). La distribución en el cuerpo del hospedador de ciertos tipos de receptores celulares explica el tropismo tisular y de hospedador de los virus.
Los tejidos y células que carecen de receptores específicos de virus determinados no son infectados por dichos viriones. La naturaleza de los receptores es variable y ciertos virus pueden tener más de un tipo de receptor.
2- Los virus complejos producen una rotura en la membrana celular del hospedador en uno de los puntos de anclaje, gracias a la presencia de algunas moléculas de enzimas hidrolíticas entre lasproteínas de la cápsida.
A través de la rotura, el tubo central inyecta el ADN vírico, quedando la cápside vacía en el exterior de la célula diana y el ácido nucleico libre en el citoplasma. La presencia de cápsidas en la superficie celular es un buen indicio de que ha sufrido una infección vírica.
Otros virus sin envoltura lipídica se introducen en la célula con cápsida y todo, lo cual puede realizarse de dos maneras:
• Por penetración directa: después de la fijación, el virus abre una brecha en la membrana y se introduce en el citoplasma.
• Por endocitosis: la membrana forma una invaginación en torno al virus, llegando a formar una vesícula que penetra en la célula.
Formada la vesícula, el virus abre una brecha en la membrana de la misma con ayuda de algunas enzimas hidrolíticas que él mismo transporta, penetrando así en el citoplasma
los virus con envoltura lipídica burlan la barrera de la membrana celular porque su cubierta lipídica se funde con la membrana, ya que son de la misma naturaleza. Esta fusión de membranas puede realizarse en dos lugares distintos:
• Fusión en la superficie celular: de manera que el virión penetra directamente en el citoplasma.
• Fusión con un lisosoma: se forma una vesícula por endocitosis, a la que se une un lisosoma para digerir la partícula introducida; entonces, la cubierta lipídica del virus se funde con la membrana del lisosoma y el virión escapa hacia el citoplasma.
3- Según la duración de la fase de eclipse, se suelen distinguir dos modalidades de ciclo infeccioso de un virus:
• Ciclo ordinario: el ácido nucleico vírico procede inmediatamente a la transcripción de su mensaje genético en los ARN necesarios para su multiplicación, y prosigue rápidamente el ciclo vital. Este tipo de ciclo es el más extendido en la naturaleza.
• Ciclo lisogénico: fue descubierto por André Lwoff en bacteriófagos. El ADN vírico se cierra por sus extremos generando un ADN circular. Este ADN se inserta en el ADN bacteriano en un lugar específico en el que la secuencia de nucleótidos bacterianos es semejante alguna región del ADN vírico.
Algunos virus que infectan células animales siguen también el ciclo lisogénico, como los papilomavirus de las verrugas y algunos retrovirus que producen algunos tipos de cáncer.
En el caso de los retrovirus, conviene recordar que el ácido nucleico es ARN monocatenario, por lo que la transcriptasa inversa ha de copiar el genoma vírico en forma de ADN antes de que pueda insertarse en el ADN celular.
4- La multiplicación del virus consta de la replicación de su material genético, de la transcripción de su mensaje en una molécula de ARN y de la traducción del mensaje para producir proteínasvíricas, tanto las que formarán parte de la cápsida como las proteínas enzimáticas necesarias para el ensamblaje de las piezas del virión y para algunas de las funciones anteriores.
Losribosomas y la mayor parte de las enzimas que los ácidos nucleicos víricos utilizan en estos procesos son los de la célula infectada.
arlin aracelis silvestre s 89308
ResponderEliminarDiapositiva 2
ADHESION
El primer paso en la infección de una célula es la adhesión o fijación a la superficie celular.
Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura.
La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula.
Las células que carecen de los receptores apropiados no son susceptibles al virus.
arlin aracelis silvestre s 89308
ResponderEliminarDiapositiva 3
EMSAMBLAJE Y MADURACION
Ensamblaje de los componentes de los virus para formar nuevas partículas víricas (proceso también llamado maduración).
Los virus desnudos realizan su ensamblaje y maduración en forma intracelular y adquieren capacidad infectante dentro de la célula. Para su salida requieren de la desintegración celular.
Los virus envueltos en el último paso al ensamblaje el virion se une con la membrana celular a la salida de la célula, mediante proteínas que se insertan en la superficie interna y exterior de la membrana.
El virion naciente brota atraves de la membrana. El ensamblaje y maduración con la expulsión atraves de la superficie celular proporcionan un mecanismo más eficaz de salida, ya que no depende de la desintegración de la célula infectada.
arlin aracelis silvestre s 89308
ResponderEliminarDiapositiva 4
PENETRACIÓN
Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus.
Virus envueltos
(A) Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática.
Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular.
(B) Entada vía endosomas en la superficie celular.
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular.
Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas.
A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma.
Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
Virus no envueltos o desnudos
Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas.
Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
arlin aracelis silvestre s 89308
ResponderEliminarDiapositiva5
SALIDA
Liberación de los nuevos virus de la célula hospedadora, que ya pueden infectar otras células.
Pueden producirse con o sin rotura de la célula, dependiendo de los virus.
Los virus desnudos salen por lisis celular lo contrario de los envueltos que salen por gemación. La distribución en el cuerpo del hospedador de ciertos tipos de receptores celulares explica el tropismo tisular y de hospedador de los virus.
Los tejidos y células que carecen de receptores específicos de virus determinados no son infectados por dichos viriones.
La naturaleza de los receptores es variable y ciertos virus pueden tener más de un tipo de receptor.
Luis E. Ametller 89066
ResponderEliminar1 Etapas de la replicacion Viral
Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada.
Luis E. Ametller 89066
ResponderEliminar2 Adhesion
El primer paso en la infección de una célula es la adhesión o fijación a la superficie celular. Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula. Las células que carecen de los receptores apropiados no son susceptibles al virus.
Luis E. Ametller 89066
ResponderEliminar3 Penetracion
Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus.
Virus envueltos
(A) Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular.
(B) Entada vía endosomas en la superficie celular.
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
Virus no envueltos o desnudos
Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
Luis E. Ametller 89066
ResponderEliminar4 Penetracion de los Bacteriografos
Los virus complejos producen una rotura en la membrana celular del hospedador en uno de los puntos de anclaje, gracias a la presencia de algunas moléculas de enzimas hidrolíticas entre las proteínas de la cápside. A través de la rotura, el tubo central inyecta el ADN vírico, quedando la cápside vacía en el exterior de la célula diana y el ácido nucleico libre en el citoplasma. La presencia de cápsidas en la superficie celular es un buen indicio de que ha sufrido una infección vírica.
Otros virus sin envoltura lipídica se introducen en la célula con cápsida y todo, lo cual puede realizarse de dos maneras:
Por penetración directa: después de la fijación, el virus abre una brecha en la membrana y se introduce en el citoplasma.
Por endocitosis: la membrana forma una invaginación en torno al virus, llegando a formar una vesícula que penetra en la célula. Formada la vesícula, el virus abre una brecha en la membrana de la misma con ayuda de algunas enzimas hidrolíticas que él mismo transporta, penetrando así en el citoplasma.
Los virus con envoltura lipídica burlan la barrera de la membrana celular porque su cubierta lipídica se funde con la membrana, ya que son de la misma naturaleza. Esta fusión de membranas puede realizarse en dos lugares distintos:
Fusión en la superficie celular: de manera que el virión penetra directamente en el citoplasma.
Fusión con un lisosoma: se forma una vesícula por endocitosis, a la que se une un lisosoma para digerir la partícula introducida; entonces, la cubierta lipídica del virus se funde con la membrana del lisosoma y el virión escapa hacia el citoplasma.
Luis E. Ametller 89066
ResponderEliminar5 Replicacion y Maduracion
Ensamblaje de los componentes de los virus para formar nuevas partículas víricas (proceso también llamado maduración).
Los virus desnudos realizan su ensamblaje y maduración en forma intracelular y adquieren capacidad infectante dentro de la célula. Para su salida requieren de la desintegración celular. Los virus envueltos en el último paso al ensamblaje el virion se une con la membrana celular a la salida de la célula, mediante proteínas que se insertan en la superficie interna y exterior de la membrana. El virion naciente brota atraves de la membrana. El ensamblaje y maduración con la expulsión atraves de la superficie celular proporcionan un mecanismo más eficaz de salida, ya que no depende de la desintegración de la célula infectada.
Luis E. Ametller 89066
ResponderEliminar6 Maduracion y salida de virus desnudos y envueltos
Después de la multiplicación del virus tiene lugar la salida de los nuevos individuos, que saldrán con capacidad de infectar nuevas células. Las principales modalidades de liberación da nombre a nuevas variantes del ciclo vital de los virus:
Infección persistente
Los nuevos virus no esperan a la muerte de la célula hospedadora para abandonarla, sino que van saliendo de la célula al mismo tiempo que se van produciendo, de manera que la célula puede seguir viva y produciendo nuevas partículas víricas. La liberación puede hacerse de dos maneras:
Los virus sin envoltura lipoproteica salen directamente, sin arrastrar ningún resto de la membrana plasmática, bien sea abriendo una brecha en la membrana, o bien aprovechando los mecanismos de exocitosis o salida de sustancias al exterior de la célula.
Luis E. Ametller 89066
ResponderEliminar7 Salida de virus envuelto
Liberación de los nuevos virus de la célula hospedadora, que ya pueden infectar otras células. Pueden producirse con o sin rotura de la célula, dependiendo de los virus. Los virus desnudos salen por lisis celular lo contrario de los envueltos que salen por gemación.
Slide 1 al 7
ResponderEliminarLa replicación viral se divide en diversas fases o etapas y son:
1) reconocimiento y fijación a la célula hospedadora, mediado por receptores específicos;
2) penetración del virus al interior de la célula mediante endocitosis o fusión de membranas;
3) pérdida de la cápside y liberación del genoma viral;
4) síntesis precoz de RNAm viral y de proteínas virales no estructurales necesarias para la replicación y la transcripción viral;
5) replicación del genoma viral;
6) síntesis tardía de RNAm viral y de proteínas estructurales;
7) modificación postranscripcional de las proteínas virales;
8) ensamblaje y formación de las partículas virales, que se puede realizar en distintas partes de la célula (núcleo, citoplasma, membrana plasmática, etc.);
9) liberación del virus mediante lisis celular, exocitosis o gemación;
10) maduración de la partícula viral (partícula infectiva)
1.- Fijación, penetración y pérdida de la cubierta (iniciación)
• El virus se fija o adsorbe a un sitio de la superficie celular que actúan como receptores específicos.
• Después entra a la célula (penetración)
• Y por último pierde la cubierta poco después de haber ingresado a la célula
2.- Síntesis de los componentes vitales (replicación)
• Inicia la fase de sinética de la replicación viral
• Se sintetizan todas las macromoléculas especificas del virus.
• Las proteínas son sintetizadas en el citoplasma
• El DNA suele replicarse en el núcleo, mientras que el ARN viral se duplica en el citoplasma (existen excepciones)
3.- Morfogénesis y liberación (fase de liberación)
• Formación de los virus descendientes
• Adquisición de la cubierta
• Muerte de la célula hospedadora (varia)
Los virus cuentan con 2 ciclos de replicación. Algunos virus se pueden replicar por lo que se conoce como ciclo Lítico; ellos inyectan a la célula huésped con su ADN, obligándola a fabricar nuevos virus, hasta que la célula huésped explota liberando los patógenos al medio.Otros virus operan de diferente forma: ellos entran e inyectan su ADN en la célula huésped pero, en lugar de tomar el control y fabricar mas virus, el ADN inyectado puede tornarse inactivo por un cierto tiempo, hasta que un apropiado evento celular dispara el proceso nuevamente. A éste ciclo se le conoce como Lisogènico.El bacteriófago, también conocido como fago, es un elemento extra cromosomal del estudio de las bacterias, en otras palabras, es un virus capaz de infectar a células eucariontes y procariontes, son capaces de replicarse por si solos, portan información genética y pueden encontrarse en casi todos los lugares pero por lo general se localizan donde están sus huéspedes obligatorios. Se reproduce en células procariontes.Esta formado por: Cabeza, cuello, cola, Vaina, placa basal, fibras de la cola y espículas.El bacteriófago también lleva a cabo un ciclo de replicación el cual se puede dividir en distintas etapas, las cuales son muy comunes a otros virus bacterianos y eucarióticos.
Replicación de los virus ADN
La mayoría de los virus ADN codifican su propia polimerasa ADN, excepto los virus ADN pequeños, como los parvovirus, papillomavirus. Esta diferencia, significa que los virus ADN pequeños necesitan células huésped en fase mitótica para suministrarles la ADN polimerasa celular
Hay 3 maneras en que los virus ADN replican su material genético:
Replicación bidireccional a partir de un substrato circular. En este caso, hay una síntesis simultánea de las cadenas ADN líder y de cierre, empezando en un origen definido de la replicación de ADN. Ejem. Los papovavirus.
Replicación desde un substrato linear. En este caso, la síntesis de las nuevas cadenas de ADN no es simultánea. Ocurre secuencialmente (primero se hace una cadena en su totalidad, luego la siguiente, etc). Un ejemplo: los adenovirus.
Replicación por medio de un intermediario ARN. Un ejemplo de este sistema son los hepaADNvirus (HBV), los cuales tienen parcialmente un genoma dsADN que debe ser convertido en un complejo ARN por la enzima transcriptasa inversa viral, durante el ciclo vital del virus.
MATRICULA-89435
ResponderEliminarEVENTOS PRINCIPALES EN LA REPLICACIÓN VIRAL
1. ADSORPCIÓN
El primer paso en la infección de una célula es la adhesión o fijación a la superficie celular. Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula. Las células que carecen de los receptores apropiados no son susceptibles al virus.
2. PENETRACIÓN
Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus.
Virus envueltos
(A) Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular. (Figura 1).
(B) Entada vía endosomas en la superficie celular (figura 2)
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
Virus no envueltos o desnudos
Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
3. PÉRDIDA DE LA CÁPSULA
El ácido nucleido debe de estar lo suficientemente expuesto como para que la replicación viral comience en esta fase. Cuando el ácido nucleico es expuesto, las partículas víricas infecciosas no pueden removerse de las células – este es el comienz de la fase de ECLIPSE – la cual perdura hasta que viriones infecciosos nuevos sean creados.
4. SÍNTESIS DE ÁCIDO NUCLEICO Y PROTEINAS VIRALES
Existen diversos mecanismos, algunos seran abarcados en capítulos posteriores.
5. ENSAMBLAJE/MADURACIÓN
Nuevas partículas víricas son ensambladas. Puede haber un estadio de maduración posterior al proceso inicial de ensamblaje.
6. LIBERACIÓN O DESCARGA
Los virus pueden ser liberados mediante lisis celular, o, si son envueltos, pueden yemar de las células. Los virus que yeman (figuras 3 y 4) no necesariamente destruyen la célula. Por tanto, algunos de estos virus son capaces de instaurar infecciones persistentes. No todas las partículas víricas liberadas son infecciosas. La proporción partículas infecciosas:partículas no-infecciosas varía según el virus y las condiciones de crecimiento.
espués de la multiplicación del virus tiene lugar la salida de los nuevos individuos, que saldrán con capacidad de infectar nuevas células. Las principales modalidades de liberación da nombre a nuevas variantes del ciclo vital de los virus:
Infección persistente[editar · editar código]
Los nuevos virus no esperan a la muerte de la célula hospedadora para abandonarla, sino que van saliendo de la célula al mismo tiempo que se van produciendo, de manera que la célula puede seguir viva y produciendo nuevas partículas víricas. La liberación puede hacerse de dos maneras:
Los virus sin envoltura lipoproteica salen directamente, sin arrastrar ningún resto de la membrana plasmática, bien sea abriendo una brecha en la membrana, o bien aprovechando los mecanismos de exocitosis o salida de sustancias al exterior de la célula.
Los virus con envoltura lipoproteica salen por gemación, es decir, se rodean de una porción de membrana plasmática que acaba separándose de la célula y constituye la cubierta lipoproteica del nuevo virus.
diana alexandra santana vasquez89309
ResponderEliminardiapositiva#1
ETAPAS DE LA REPLICACIÓN VIRAL
1- El primer paso en la infección viral es la absorción a la membrana de la célula susceptible por medio de la adhesión de ligandos virales (proteínas de la capside o glucoproteínas de las espículas, por ejemplo a receptores superficiales de la célula). La distribución en el cuerpo del hospedador de ciertos tipos de receptores celulares explica el tropismo tisular y de hospedador de los virus. Los tejidos y células que carecen de receptores específicos de virus determinados no son infectados por dichos viriones. La naturaleza de los receptores es variable y ciertos virus pueden tener más de un tipo de receptor.
2- Los virus complejos producen una rotura en la membrana celular del hospedador en uno de los puntos de anclaje, gracias a la presencia de algunas moléculas de enzimas hidrolíticas entre lasproteínas de la cápsida. A través de la rotura, el tubo central inyecta el ADN vírico, quedando la cápside vacía en el exterior de la célula diana y el ácido nucleico libre en el citoplasma. La presencia de cápsidas en la superficie celular es un buen indicio de que ha sufrido una infección vírica.
Otros virus sin envoltura lipídica se introducen en la célula con cápsida y todo, lo cual puede realizarse de dos maneras:
• Por penetración directa: después de la fijación, el virus abre una brecha en la membrana y se introduce en el citoplasma.
• Por endocitosis: la membrana forma una invaginación en torno al virus, llegando a formar una vesícula que penetra en la célula. Formada la vesícula, el virus abre una brecha en la membrana de la misma con ayuda de algunas enzimas hidrolíticas que él mismo transporta, penetrando así en el citoplasma.
Los virus con envoltura lipídica burlan la barrera de la membrana celular porque su cubierta lipídica se funde con la membrana, ya que son de la misma naturaleza. Esta fusión de membranas puede realizarse en dos lugares distintos:
• Fusión en la superficie celular: de manera que el virión penetra directamente en el citoplasma.
• Fusión con un lisosoma: se forma una vesícula por endocitosis, a la que se une un lisosoma para digerir la partícula introducida; entonces, la cubierta lipídica del virus se funde con la membrana del lisosoma y el virión escapa hacia el citoplasma.
3- Según la duración de la fase de eclipse, se suelen distinguir dos modalidades de ciclo infeccioso de un virus:
• Ciclo ordinario: el ácido nucleico vírico procede inmediatamente a la transcripción de su mensaje genético en los ARN necesarios para su multiplicación, y prosigue rápidamente el ciclo vital. Este tipo de ciclo es el más extendido en la naturaleza.
• Ciclo lisogénico: fue descubierto por André Lwoff en bacteriófagos. El ADN vírico se cierra por sus extremos generando un ADN circular. Este ADN se inserta en el ADN bacteriano en un lugar específico en el que la secuencia de nucleótidos bacterianos es semejante alguna región del ADN vírico.
Algunos virus que infectan células animales siguen también el ciclo lisogénico, como los papilomavirus de las verrugas y algunos retrovirus que producen algunos tipos de cáncer.
En el caso de los retrovirus, conviene recordar que el ácido nucleico es ARN monocatenario, por lo que la transcriptasa inversa ha de copiar el genoma vírico en forma de ADN antes de que pueda insertarse en el ADN celular.
4- La multiplicación del virus consta de la replicación de su material genético, de la transcripción de su mensaje en una molécula de ARN y de la traducción del mensaje para producir proteínasvíricas, tanto las que formarán parte de la cápsida como las proteínas enzimáticas necesarias para el ensamblaje de las piezas del virión y para algunas de las funciones anteriores. Losribosomas y la mayor parte de las enzimas que los ácidos nucleicos víricos utilizan en estos procesos son los de la célula infectada.
diana alexandra santana vasquez89309
ResponderEliminardiapositva#2
Adhesion
El primer paso en la infección de una célula es la adhesión o fijación a la superficie celular. Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula.
Las células que carecen de los receptores apropiados no son susceptibles al virus.
ETAPAS DE LA REPLICACION VIRAL
ResponderEliminarSlide 1: Etapas de la replicación viral 2012-0593
El ciclo de replicación de los virus es el término utilizado para describir el ciclo de reproducción de los virus. Este consta generalmente de las siguientes fases: fijación y entrada en la célula, eclipse, multiplicación y liberación del virus. Los virus son formas acelulares que no pueden reproducirse por si mismos en el exterior; para ello requieren introducirse en células donde controlarán sus mecanismos reproductivos
Slide 2: Absorcion 2012-0593
El primer paso en la infección de una célula es la adhesión o fijación a la superficie celular. Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula. Las células que carecen de los receptores apropiados no son susceptibles al virus.
Slide 3: Penetracion 2012-0593
La penetración a la célula blanco ocurre normalmente en un tiempo muy breve luego de la unión del virus al receptor en la superficie celular. A diferencia del proceso anterior, el ingreso a la célula es generalmente un proceso dependiente de energía, esto significa que la célula debe estar metabolicamente activa .
En este paso hay tres mecanismos involucrados:
1) TRANSLOCACIÓN: La partícula viral atraviesa la membrana citoplasmática. Este proceso es poco frecuente en los virus y no está completamente entendido. Puede estar mediado por proteínas de la cápside viral y receptores específicos de membrana
2) ENDOCITOSIS: El virus ingresa en vacuolas intracelulares. Este es probablemente el mecanismo de ingreso más común en los virus. No se necesita de la presencia de proteínas virales específicas -excepto de aquellas involucradas en la unión al receptor- pero si es necesaria la formación de vesículas endocíticas en la membrana celular y la posterior internalización de estas. Este mecanismo involucra un cambio de pH en el endosoma para liberar posteriormente el genoma del virus dentro del citoplasma.
3) FUSIÓN: Sólo ocurre con los virus envueltos. En este proceso ocurre una fusión de la envoltura viral con la membrana celular, ya sea directamente en la superficie celular o en una vesícula citoplasmática luego de la endocitosis. La fusión requiere de la presencia de una proteína de fusión específica en la envoltura del virus -por ejemplo la hemaglutinina en el virus de la influenza o las glucoproteínas de transmembrana en los retrovirus-. Estas proteínas promueven la unión de las membranas del virus y de la célula, esto resulta en la liberación de la nucleocápside directamente en el citoplasma.
Este proceso no es dependiente pH.
El proceso de endocitosis es casi universal en las células animales; se forman vesículas en la membrana plasmática, las que posteriormente son englobadas e internalizadas al citoplasma. La vida media de estas vesículas es muy breve, en pocos segundos la mayoría se fusionan con endosomas liberando su contenido en esas vesículas más grandes. Hasta este momento cualquier virus contenido en estas estructuras, aún está separado del citoplasma por una bicapa lipídica , por lo tanto no se puede decir estrictamente que ha ingresado en la célula. Posteriormente, los endosomas se fusionan con los lisosomas, el ambiente interno se vuelve progresivamente más hostil, se acidifica en la medida que el pH desciende y aumenta la concentración de enzimas degradativas . Esto significa que el virus debe liberarse de la vesícula e ingresar al citoplasma antes de ser degradado. Hay una serie de mecanismos por los cuales esto ocurre, incluyendo la fusión de membranas. La liberación de la partícula viral desde el endosoma y su pasaje al citoplasma están conectados estrechamente -y a menudo imposible de separar- del proceso de liberación de las estructuras, liberación del genoma -.
diana alexandra santana vasquez89309
ResponderEliminardiapositiva#3
PENETRACIÓN
Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus.
Virus envueltos
(A) Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular.
(B) Entada vía endosomas en la superficie celular.
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
Virus no envueltos o desnudos
Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
Slide 4: Penetracion Bacteriofago 2012-0593
ResponderEliminarLa primera etapa de la infección viral es la interacción de un virión con un sitio receptor específica de la superficie celular. Las moléculas receptoras difieren para los distintos virus, y son proteínas en algunos casos (por ejemplo, picornavirus) y oligosacáridos en otros (por ejemplo, ortovirus y paramixovirus). La presencia o la ausencia de receptores desempeña una función determinante de primer orden en el tropismo celular y la patogenia viral; por ejemplo, el poliovirus es capaz de fijarse sólo en las células del sistema nervioso central y las vías digestivas de los primates. Se cree que la fijación en receptores refleja homologías configurativas fortuitas entre la estructura de la superficie del virión y un componente de la superficie celular. Por ejemplo, el virus de la inmunodeficiencia humana (HIV) se une al receptor CD4 en las células del sistema inmunitario y se cree que el virus de la rabia interactúa con los receptores de acetilcolina y que el virus de Epstein-Barr reconoce al receptor para el tercer componente del complemento en las células B. Es probable que cada célula susceptible posea por lo menos 100,000 sitios receptores para un virus dado
Slide 5: Replicacion y Maduracion 2012-0593
Este es el estadío del ciclo replicativo en el cual el virus se hace infeccioso. La maduración generalmente incluye cambios estructurales en la partícula viral, estos cambios pueden ser el resultado de cortes específicos de proteínas de la cápside para formar productos maduros, o de cambios conformacionales en las proteínas durante el ensamble. Estos eventos conducen frecuentemente a cambios estructurales sustanciales en la cápside, por ej. las diferencias de antigenicidad entre la partícula viral incompleta y la madura.
Las proteasas virales frecuentemente están involucradas en la maduración, aunque en algunos casos pueden actuar enzimas celulares o una mezcla de enzimas celulares y virales. Ciertamente hay un cierto riesgo en la participación de enzimas proteolíticas celulares en este proceso, dado que la falta de especificidad sobre el sustrato podría ocasionar fácilmente la completa degradación de las proteínas de la cápside.
Sin embargo las proteasas codificadas por el virus, en general son altamente específicas para determinadas secuencias de aminoácidos y estructuras; frecuentemente sólo cortan una unión peptídica en particular en una estructura proteica grande y compleja como la cápside. Además, en cierta forma son controladas al ser empaquetadas dentro de la partícula viral durante el ensamble y sólo se activan cuando toman contacto con las secuencias específicas para la conformación de la cápside, por ejemplo al localizarse en un sitio hidrofóbico o por cambios de pH o concentración de determinados iones dentro de la cápside. Las proteasas de los retrovirus son un buen ejemplo de enzimas involucradas en la maduración, que están bajo este tipo de control. El core de los retrovirus está compuesto por proteínas codificadas por el gen gag y la proteasa es empaquetada en el core antes de la liberación de la célula por gemación. En algún momento del proceso de gemación, la proteasa corta los precursores de la proteína gag en productos maduros -proteínas de la càpside, nucleocàpside y matriz de la partícula viral madura-. Para algunos virus el ensamble y la maduración ocurren dentro de la célula y son procesos inseparables, mientras que para otros la maduración puede ocurrir sólo después de la liberación de la partícula viral. De todos modos, y en cualquiera de los casos, el proceso de maduración prepara al virus para la infección de nuevas células. En resumen, la replicación de los virus implica tres procesos básicos:
a) El inicio de la infección.
b) La replicación y expresión del genoma.
c) La liberación del virión maduro desde la célula infectada
Slide 6: Maduracion y Lberacion 2012-0593
ResponderEliminarLos virus en general pueden liberarse de la célula mediante dos tipos de mecanismos: lisis y gemación.
Para los virus líticos -la mayoría de los virus desnudos-, la liberación es un proceso simple: la célula infectada se rompe y libera las partículas virales. En el caso de los virus que se liberan por gemación, estos adquieren su envoltura lipídica y las proteínas asociadas en el momento en que pasan a través de la membrana plasmática o de vesículas intracelulares. La liberación de los virus por esta vía puede resultar altamente perjudicial para la célula -por ej.en el caso de paramixovirus, rabdovirus, togavirus- o no -por ej. retrovirus-; de todos modos en cualquiera de los casos el proceso es controlado por el virus: la interacción física de las proteínas de la cápside sobre la superficie interna de la membrana celular fuerza a la partícula viral a pasar a través de dicha membrana.
El ensamble, la maduración y liberación son generalmente procesos simultáneos en los virus que se liberan por gemación. El tipo de membrana a través de la cual geman los virus depende del tipo de virus, por ej. puede ser a través de la membrana celular como en el caso de los retrovirus y togavirus, a través de la membrana citoplasmática -ortomixovirus, coronavirus, bunyavirus, rabdovirus, hepadnavirus- o a través de la membrana nuclear como en los herpesvirus.
Slide 7: Salida de un virus envuelto 2012-0593
Se encuentra la variación más amplia en las estrategias de la expresión genética entre los virus que contienen ARN. Algunos viriones poseen polimerasas (ortomixovirus, reovirus); otros sistemas emplean mensajes subgenómicos generados a veces por empalme (ortomixovirus, retrovirus); por último, algunos virus sintetizan grandes precursores poliproteínicos que se someten a procesamiento y segmentación para generar los productos génicos finales (picornavirus, retrovirus).Los sitios intracelulares en los que ocurren los diferentes acontecimientos de la replicación viral, varían de un grupo a otro. Es posible plantear algunas generalizaciones. La proteína viral se sintetiza en el citoplasma, sobre los polirribosomas compuestos por ARNm específico del virus y ribosomas de la célula huésped. El ADN viral suele replicarse en el núcleo. El ARN genómico viral se duplica generalmente en el citoplasma celular, aunque hay excepciones
diana alexandra santana vasquez89309
ResponderEliminardiapositiva#4
La primera etapa de la infección viral es la interacción de un virión con un sitio receptor específica de la superficie celular.
Las moléculas receptoras difieren para los distintos virus, y son proteínas en algunos casos (por ejemplo, picornavirus) y oligosacáridos en otros (por ejemplo, ortovirus y paramixovirus).
La presencia o la ausencia de receptores desempeña una función determinante de primer orden en el tropismo celular y la patogenia viral; por ejemplo, el poliovirus es capaz de fijarse sólo en las células del sistema nervioso central y las vías digestivas de los primates. Se cree que la fijación en receptores refleja homologías configurativas fortuitas entre la estructura de la superficie del virión y un componente de la superficie celular. Por ejemplo, el virus de la inmunodeficiencia humana (HIV) se une al receptor CD4 en las células del sistema inmunitario y se cree que el virus de la rabia interactúa con los receptores de acetilcolina y que el virus de Epstein-Barr reconoce al receptor para el tercer componente del complemento en las células B.
Es probable que cada célula susceptible posea por lo menos 100,000 sitios receptores para un virus dado.
diana alexandra santana vasquez89309
ResponderEliminardiapositiva#5
mediante Después de la fijación, la partícula viral entra en la célula. Esta etapa se conoce como penetración, viropexia o fagocitosis.
En algunos sistemas, se realiza por endocitosis mediada por receptores, con captación de las partículas virales ingeridas dentro de endosomas.
Están menos claros los detalles de la penetración en otros sistemas. Ocurre perdida de la cubierta de manera concomitante con la penetración o poco después de la misma. La pérdida de la cubierta, o descubrimiento, es la separación física del ácido nucleico viral (o, en algunos casos, de las nucleocápsides inteARNs) a partir de los componentes estructurales externos del virión. En este momento se pierde la infecciosidad del virus original.
Los virus son los únicos agentes infecciosos en los que la disolución del agente infectante es una etapa obligatoria de la vía de la replicación
diana alexandra santana vasquez89309
ResponderEliminardiapositiva#6
Maduracion y salida de virus desnudos y envueltos
Después de la multiplicación del virus tiene lugar la salida de los nuevos individuos, que saldrán con capacidad de infectar nuevas células. Las principales modalidades de liberación da nombre a nuevas variantes del ciclo vital de los virus:
Infección persistente
Los nuevos virus no esperan a la muerte de la célula hospedadora para abandonarla, sino que van saliendo de la célula al mismo tiempo que se van produciendo, de manera que la célula puede seguir viva y produciendo nuevas partículas víricas
La liberación puede hacerse de dos maneras:
Los virus sin envoltura lipoproteica salen directamente, sin arrastrar ningún resto de la membrana plasmática, bien sea abriendo una brecha en la membrana, o bien aprovechando los mecanismos de exocitosis o salida de sustancias al exterior de la célula.
diana alexandra santana vasquez89309
ResponderEliminardiapositiva#7
SALIDA
Liberación de los nuevos virus de la célula hospedadora, que ya pueden infectar otras células. Pueden producirse con o sin rotura de la célula, dependiendo de los virus. Los virus desnudos salen por lisis celular lo contrario de los envueltos que salen por gemación.
-todo Se encuentra la variación más amplia en las estrategias de la expresión genética entre los virus que contienen ARN. Algunos viriones poseen polimerasas (ortomixovirus, reovirus); otros sistemas emplean mensajes subgenómicos generados a veces por empalme (ortomixovirus, retrovirus); por último, algunos virus sintetizan grandes precursores poliproteínicos que se someten a procesamiento y segmentación para generar los productos génicos finales (picornavirus, retrovirus).
La extensión a la que las enzimas específicas del virus participan en estos procesos varía de un grupo a otro. En general, los virus de mayor tamaño (herpesvirus, poxvirus) son más independientes de las funciones celulares que los virus de menor tamaño. Esta es una razón de que los virus de mayor tamaño sean más sensibles a la quimioterapia antiviral, porque se dispone de más procesos específicos de los virus como blancos de la acción farmacológica.
Los sitios intracelulares en los que ocurren los diferentes acontecimientos de la replicación viral, varían de un grupo a otro. Es posible plantear algunas generalizaciones. La proteína viral se sintetiza en el citoplasma, sobre los polirribosomas compuestos por ARNm específico del virus y ribosomas de la célula huésped. El ADN viral suele replicarse en el núcleo. El ARN genómico viral se duplica generalmente en el citoplasma celular, aunque hay excepciones
2011-0029 José Luis González Salas
ResponderEliminarDiapositiba1
Etapas de la replicación viral
La replicación es el proceso en el cual se copia el ADN. Este proceso es semiconservativo y bidireccional. Funciona igual, tanto en procariontes como en eucarionte, salvo algunos cambio, principalmente de las proteínas que participan. Se dieron muchas hipótesis sobre como se dupllicaba el ADN hasta que Watson y Crick propusieron la hipótesis semiconservativa, según la cual, las nuevas moléculas de ADN formadas a partir de otra antigua, tienen una hebra antigua y otra nueva. La replicación ocurre en 3 etapas: La iniciación La elongación La terminación.
Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada.
El ciclo de replicación de los virus es el término utilizado para describir el ciclo de reproducción de los virus. Este consta generalmente de las siguientes fases: fijación y entrada en la célula, eclipse, multiplicación y liberación del virus. Los virus son formas acelulares que no pueden reproducirse por si mismos en el exterior; para ello requieren introducirse en células donde controlarán sus mecanismos reproductivos.
2011-0029 José Luis González Salas
ResponderEliminarDiapositiva 2
ADHESION
El primer paso en la infección de una célula es la adhesión o fijación a la superficie celular. Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula. Las células que carecen de los receptores apropiados no son susceptibles al virus.
2011-0029 José Luis González Salas
ResponderEliminarDiapositiva 3
PENETRACIÓN
Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus.
Virus envueltos
(A) Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular.
(B) Entada vía endosomas en la superficie celular.
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
Virus no envueltos o desnudos
Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
2011-0029 José Luis González Salas
ResponderEliminarDiapositiva 4
EMSAMBLAJE Y MADURACION
Ensamblaje de los componentes de los virus para formar nuevas partículas víricas (proceso también llamado maduración).
Los virus desnudos realizan su ensamblaje y maduración en forma intracelular y adquieren capacidad infectante dentro de la célula. Para su salida requieren de la desintegración celular. Los virus envueltos en el último paso al ensamblaje el virion se une con la membrana celular a la salida de la célula, mediante proteínas que se insertan en la superficie interna y exterior de la membrana. El virion naciente brota atraves de la membrana. El ensamblaje y maduración con la expulsión atraves de la superficie celular proporcionan un mecanismo más eficaz de salida, ya que no depende de la desintegración de la célula infectada.
2011-0029 José Luis González Salas
ResponderEliminarDiapositiva5
SALIDA
Liberación de los nuevos virus de la célula hospedadora, que ya pueden infectar otras células. Pueden producirse con o sin rotura de la célula, dependiendo de los virus. Los virus desnudos salen por lisis celular lo contrario de los envueltos que salen por gemación. La distribución en el cuerpo del hospedador de ciertos tipos de receptores celulares explica el tropismo tisular y de hospedador de los virus. Los tejidos y células que carecen de receptores específicos de virus determinados no son infectados por dichos viriones. La naturaleza de los receptores es variable y ciertos virus pueden tener más de un tipo de receptor.
2011-0029 José Luis González Salas
ResponderEliminarDiapositiva 6
Maduracion y salida de virus desnudos y envueltos
Después de la multiplicación del virus tiene lugar la salida de los nuevos individuos, que saldrán con capacidad de infectar nuevas células. Las principales modalidades de liberación da nombre a nuevas variantes del ciclo vital de los virus:
Infección persistente
Los nuevos virus no esperan a la muerte de la célula hospedadora para abandonarla, sino que van saliendo de la célula al mismo tiempo que se van produciendo, de manera que la célula puede seguir viva y produciendo nuevas partículas víricas
La liberación puede hacerse de dos maneras:
Los virus sin envoltura lipoproteica salen directamente, sin arrastrar ningún resto de la membrana plasmática, bien sea abriendo una brecha en la membrana, o bien aprovechando los mecanismos de exocitosis o salida de sustancias al exterior de la célula.
2011-0029 José Luis González Salas
ResponderEliminarDiapositiva 7
SALIDA
Liberación de los nuevos virus de la célula hospedadora, que ya pueden infectar otras células. Pueden producirse con o sin rotura de la célula, dependiendo de los virus. Los virus desnudos salen por lisis celular lo contrario de los envueltos que salen por gemación.
-todo Se encuentra la variación más amplia en las estrategias de la expresión genética entre los virus que contienen ARN. Algunos viriones poseen polimerasas (ortomixovirus, reovirus); otros sistemas emplean mensajes subgenómicos generados a veces por empalme (ortomixovirus, retrovirus); por último, algunos virus sintetizan grandes precursores poliproteínicos que se someten a procesamiento y segmentación para generar los productos génicos finales (picornavirus, retrovirus).
La extensión a la que las enzimas específicas del virus participan en estos procesos varía de un grupo a otro. En general, los virus de mayor tamaño (herpesvirus, poxvirus) son más independientes de las funciones celulares que los virus de menor tamaño. Esta es una razón de que los virus de mayor tamaño sean más sensibles a la quimioterapia antiviral, porque se dispone de más procesos específicos de los virus como blancos de la acción farmacológica.
Los sitios intracelulares en los que ocurren los diferentes acontecimientos de la replicación viral, varían de un grupo a otro. Es posible plantear algunas generalizaciones. La proteína viral se sintetiza en el citoplasma, sobre los polirribosomas compuestos por ARNm específico del virus y ribosomas de la célula huésped. El ADN viral suele replicarse en el núcleo. El ARN genómico viral se duplica generalmente en el citoplasma celular, aunque hay excepciones
89693 - Fabiano Lima das Gracas
ResponderEliminarVirologia – Grupo Lunes 10-12pm
1-Etapas de la Replicacion Viral:
Ciclo de los virus
• La adhesión o adsorción es una unión específica entre proteínas de la cápside vírica y receptores específicos de la superficie celular del huésped, pero algunos bacteriófagos también son capaces de adherirse a los flagelos, vellosidades (pili) o cápsulas presentes en la superficie de la bacteria hospedera.
• La penetración sigue a la adhesión; los virus se introducen en la célula huésped mediante endocitosis mediada por receptores o por fusión de membrana. Esto recibe a menudo el nombre de penetración vírica
• El despojo es el proceso en que la cápside vírica es degradada por enzimas virales o del huésped, liberando así el ácido nucleico del genoma vírico.
• La replicación implica la síntesis de ARN mensajero (ARNm) vírico en todos los virus con rasgos de ARN positivos, la síntesis de proteínas víricas, el ensamblaje de proteínas víricas y la replicación del genoma viral
• Tras el ensamblaje de partículas víricas, a menudo se produce una modificación postraduccional de las proteínas víricas.
89693 - Fabiano Lima das Gracas
ResponderEliminarVirologia – Grupo Lunes 10-12pm
2-Adhesion:
El primer paso en la infección de una célula es la adhesión o fijación a la superficie celular. Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula. Las células que carecen de los receptores apropiados no son susceptibles al virus.
89693 - Fabiano Lima das Gracas
ResponderEliminarVirologia – Grupo Lunes 10-12pm
3-Penetracion:
Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus.
Virus envueltos
• Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular.
• Entada vía endosomas en la superficie celular
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
Virus no envueltos o desnudos
Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
89693 - Fabiano Lima das Gracas
ResponderEliminarVirologia – Grupo Lunes 10-12pm
4-Penetracion de los Bacteriofagos:
Los bacteriófagos (también llamados fagos), son virus que infectan exclusivamente a bacterias.
Para entrar en una célula, los fagos se acoplan a receptores específicos en la superficie de la bacteria, que pueden ser lipopolisacáridos, ácidos teicoicos, proteínas o incluso flagelos. Por ello, cada fago solo podrá infectar ciertas bacterias según sus receptores. Puesto que los fagos no son móviles, dependen de encuentros al azar con los receptores adecuados en solución para poder infectar una bacteria.
Parece que los bacteriófagos presentan una especie de jeringa mediante la cual introducen su material genético en el interior de la célula. Tras el reconocimiento del receptor adecuado, la cola y cuello del fago se contraen, quedando así el fago acoplado a la superficie celular. El material genético puede ser ahora introducido a través de la membrana o bien simplemente depositado sobre la superficie. No se descarta que pueda haber fagos con otros métodos diferentes para introducir su material genético en la célula.
89693 - Fabiano Lima das Gracas
ResponderEliminarVirologia – Grupo Lunes 10-12pm
5-Replicacion y Maduracion:
Los genomas virales recién sintetizados y los polipéptidos de la cápside se ensamblan para formar los virus descendientes. Las cápsides icosaédricas pueden condensarse en ausencia de ácido nucleico, en tanto que las nucleocápsides de los virus de simetría helicoidal no pueden formarse sin ARN viral. No existen mecanismos especiales para la liberación de virus carentes de cubierta; las células infectadas acaban por experimentar lisis y liberar las partículas virales.
Los virus cubiertos maduran por un proceso de gemación. Se insertan glucoproteínas de cubierta específica del virus en las membranas celulares y, a continuación, las nucleocápsides hacen gemación a través de la membrana a nivel de estos sitios modificados y, al hacerlo así, adquieren su cubierta. La gemación ocurre a menudo a nivel de la membrana plasmática y puede abarcar a otras membranas de la célula. Los virus cubiertos no son infecciosos hasta que han adquirido sus cubiertas. Por lo tanto, no se acumulan de manera típica viriones descendientes infecciosos en el interior de la célula infectada.
La maduración viral es a veces un proceso ineficaz. Pueden acumularse cantidades excesivas de componentes virales y participar en la formación de cuerpos de inclusión en la célula Como resultado de los profundos efectos dañinos de la replicación viral, se desarrollan por último efectos citopáticos celulares y la célula muere. Sin embargo, hay casos en que el virus no lesiona a la célula y se producen infecciones persistentes y prolongadas.
89693 - Fabiano Lima das Gracas
ResponderEliminarVirologia – Grupo Lunes 10-12pm
6-Maduracion y Salida de Virus Desnudos y Envueltos:
Virus envueltos
• Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular.
• Entada vía endosomas en la superficie celular
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
Virus no envueltos o desnudos
Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
89693 - Fabiano Lima das Gracas
ResponderEliminarVirologia – Grupo Lunes 10-12pm
7-Salida de un Virus Envuelto:
Los virus pueden ser liberados mediante lisis celular, o, si son envueltos, pueden yemar de las células. Los virus que yeman no necesariamente destruyen la célula. Por tanto, algunos de estos virus son capaces de instaurar infecciones persistentes. No todas las partículas víricas liberadas son infecciosas. La proporción partículas infecciosas:partículas no-infecciosas varía según el virus y las condiciones de crecimiento.
88977
ResponderEliminarETAPAS DE REPLICACION VIRAL
El ciclo de replicación de los virus es el término utilizado para describir el ciclo de reproducción de los virus. Este consta generalmente de las siguientes fases: fijación y entrada en la célula, eclipse, multiplicación y liberación del virus. Los virus son formas acelulares que no pueden reproducirse por si mismos en el exterior; para ello requieren introducirse en células donde controlarán sus mecanismos
88977
ResponderEliminarADHESION
El primer paso en la infección de una célula es la adhesión o fijación a la superficie celular. Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula. Las células que carecen de los receptores apropiados no son susceptibles al virus.
88977
ResponderEliminarPENETRACIÓN
Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus.
Virus envueltos
(A) Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular.
(B) Entada vía endosomas en la superficie celular.
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
Virus no envueltos o desnudos
Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
88977
ResponderEliminarPENETRACION DE UN BACTERIOFAGO
ciclo litico de un bacteriófago.formado por acido nuceico ADN y capsida compleja.
fijacion o adsorcion: se fijan inicialmente a traves de las puntas de las fibras cudales mediante enlaces quimicos y posterormente de forma mecanica al clavar las pespinas basales en la pared bacteriana.
penetracion: el bacteriofago mediante enzimas lisozimas situadas en su placa basal perrfora la pared cecular y luego contrae la vaina e introduce el eje tubular de modo que el adn pasa directamente al citoplasma
88977
ResponderEliminarREPLICACION Y MADURACION
Ensamblaje de los componentes de los virus para formar nuevas partículas víricas (proceso también llamado maduración).
Los virus desnudos realizan su ensamblaje y maduración en forma intracelular y adquieren capacidad infectante dentro de la célula. Para su salida requieren de la desintegración celular. Los virus envueltos en el último paso al ensamblaje el virion se une con la membrana celular a la salida de la célula, mediante proteínas que se insertan en la superficie interna y exterior de la membrana. El virion naciente brota atraves de la membrana. El ensamblaje y maduración con la expulsión atraves de la superficie celular proporcionan un mecanismo más eficaz de salida, ya que no depende de la desintegración de la célula infectada
Salida de virus envuelto
ResponderEliminarLiberación de los nuevos virus de la célula hospedadora, que ya pueden infectar otras células. Pueden producirse con o sin rotura de la célula, dependiendo de los virus. Los virus desnudos salen por lisis celular lo contrario de los envueltos que salen por gemación.Después de la multiplicación del virus tiene lugar la salida de los nuevos individuos, que saldrán con capacidad de infectar nuevas células. Las principales modalidades de liberación da nombre a nuevas variantes del ciclo vital de los virus:
Christopher Robles
ResponderEliminar2011-0692
Etapas de la replicacion viral
Slide 1 Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada.
La única característica de la multiplicación viral consiste en que, poco después de su interacción con una célula huésped, el virión infectante se desintegra y pierde la infecciosidad que se puede medir. Esta fase del ciclo celular se llama periodo de eclipse; su duración varía dependiendo del virus y de la célula huésped en particular, y termina con la formación de las primeras partículas virales descendientes infecciosas. El periodo de eclipse es, en realidad, de intensa actividad sintética, puesto que la célula se redirige hacia la satisfacción de las necesidades del "pirata" viral. En algunos casos, tan pronto como el ácido nucleico viral penetra a la célula huésped, el metabolismo celular es recanalizado exclusivamente hacia la síntesis de nuevas partículas virales. En otros casos, los procesos metabólicos de la célula huésped no se alteran significativamente, aunque la célula sintetiza proteínas virales y ácidos nucleicos.
Christopher Robles
ResponderEliminar2011-0692
Slide 2, Los virus han desarrollado diversas estrategias para lograr la multiplicación en las células huéspedes parasitadas por ellos. Aunque los detalles varían de un grupo a otro, el patrón general de los ciclos de replicación es semejante. La primera etapa de la infección viral es la interacción de un virión con un sitio receptor específica de la superficie celular. Las moléculas receptoras difieren para los distintos virus, y son proteínas en algunos casos (por ejemplo, picornavirus) y oligosacáridos en otros (por ejemplo, ortovirus y paramixovirus). La presencia o la ausencia de receptores desempeña una función determinante de primer orden en el tropismo celular y la patogenia viral; por ejemplo, el poliovirus es capaz de fijarse sólo en las células del sistema nervioso central y las vías digestivas de los primates. Se cree que la fijación en receptores refleja homologías configurativas fortuitas entre la estructura de la superficie del virión y un componente de la superficie celular. Por ejemplo, el virus de la inmunodeficiencia humana (HIV) se une al receptor CD4 en las células del sistema inmunitario y se cree que el virus de la rabia interactúa con los receptores de acetilcolina y que el virus de Epstein-Barr reconoce al receptor para el tercer componente del complemento en las células B. Es probable que cada célula susceptible posea por lo menos 100,000 sitios receptores para un virus dado.
Christopher Robles
ResponderEliminar2011-0692
Slide 3-4, Esta etapa se conoce como penetración, viropexia o fagocitosis. En algunos sistemas, se realiza por endocitosis mediada por receptores, con captación de las partículas virales ingeridas dentro de endosomas. Están menos claros los detalles de la penetración en otros sistemas. Ocurre perdida de la cubierta de manera concomitante con la penetración o poco después de la misma. La pérdida de la cubierta, o descubrimiento, es la separación física del ácido nucleico viral (o, en algunos casos, de las nucleocápsides inteARNs) a partir de los componentes estructurales externos del virión. En este momento se pierde la infecciosidad del virus original. Los virus son los únicos agentes infecciosos en los que la disolución del agente infectante es una etapa obligatoria de la vía de la replicación. Poco después de quedar descubierto el genoma viral, se inicia la fase sintética del ciclo de replicación del virus. El aspecto esencial de la replicación viral consiste en que deben transcribirse los ARNm específicos del ácido nucleico viral para que se exprese y duplique con éxito la información genética. Una vez que se logra esta etapa, los virus recurren a los componentes de la célula para traducir el ARNm. Diversas clases de virus emplean vías diferentes para sintetizar los ARNm según la estructura del ácido nucleico viral. Algunos virus (por ejemplo, rhabdovirus, ortomixovirus y paramixovirus) poseen ARN polimerasas que sintetizan a los ARNm. Los virus ARN de este tipo se llaman virus de tira negativa (de sentido negativo), puesto que su genoma de ARN de una sola tira es complementario del ARNm, el cual se designa por acuerdo general como de tira positiva (o de sentido positivo). Durante la replicación viral se sintetizan todas las macromoléculas especificadas del virus en una secuencia muy organizada. En algunas infecciones por virus, notablemente las producidas por virus de doble tira que contienen ADN, se sintetizan proteínas virales tempranas poco después de la infección y aparecen proteínas tardías sólo durante la fase tardía del proceso infeccioso, una vez que ha ocurrido la síntesis del ADN viral. Pueden activarse o no genes tempranos cuando se elaboran los productos tardíos. En contraste, se expresa al mismo tiempo la mayor parte de la información genética de los virus que contienen ARN o toda ella. Además de estos controles temporales, existen también controles cuantitativos puesto que no se elaboran todas las proteínas virales en las mismas cantidades. Las proteínas específicas del virus pueden regular la extensión de la transcripción del genoma o la traducción del ARNm viral.
Christopher Robles
ResponderEliminar2011-0692
Slide 5-6, Se encuentra la variación más amplia en las estrategias de la expresión genética entre los virus que contienen ARN. Algunos viriones poseen polimerasas (ortomixovirus, reovirus); otros sistemas emplean mensajes subgenómicos generados a veces por empalme (ortomixovirus, retrovirus); por último, algunos virus sintetizan grandes precursores poliproteínicos que se someten a procesamiento y segmentación para generar los productos génicos finales (picornavirus, retrovirus). La extensión a la que las enzimas específicas del virus participan en estos procesos varía de un grupo a otro. En general, los virus de mayor tamaño (herpesvirus, poxvirus) son más independientes de las funciones celulares que los virus de menor tamaño. Esta es una razón de que los virus de mayor tamaño sean más sensibles a la quimioterapia antiviral, porque se dispone de más procesos específicos de los virus como blancos de la acción farmacológica. Los sitios intracelulares en los que ocurren los diferentes acontecimientos de la replicación viral, varían de un grupo a otro. Es posible plantear algunas generalizaciones. La proteína viral se sintetiza en el citoplasma, sobre los polirribosomas compuestos por ARNm específico del virus y ribosomas de la célula huésped. El ADN viral suele replicarse en el núcleo. El ARN genómico viral se duplica generalmente en el citoplasma celular, aunque hay excepciones.
Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada
ResponderEliminarLa única característica de la multiplicación viral consiste en que, poco después de su interacción con una célula huésped, el virión infectante se desintegra y pierde la infecciosidad que se puede medir. Esta fase del ciclo celular se llama periodo de eclipse; su duración varía dependiendo del virus y de la célula huésped en particular, y termina con la formación de las primeras partículas virales descendientes infecciosas. El periodo de eclipse es, en realidad, de intensa actividad sintética, puesto que la célula se redirige hacia la satisfacción de las necesidades del "pirata" viral. En algunos casos, tan pronto como el ácido nucleico viral penetra a la célula huésped, el metabolismo celular es recanalizado exclusivamente hacia la síntesis de nuevas partículas virales. En otros casos, los procesos metabólicos de la célula huésped no se alteran significativamente, aunque la célula sintetiza proteínas virales y ácidos nucleicos.
La primera etapa de la infección viral es la interacción de un virión con un sitio receptor específica de la superficie celular. Las moléculas receptoras difieren para los distintos virus, y son proteínas en algunos casos (por ejemplo, picornavirus) y oligosacáridos en otros (por ejemplo, ortovirus y paramixovirus). La presencia o la ausencia de receptores desempeña una función determinante de primer orden en el tropismo celular y la patogenia viral; por ejemplo, el poliovirus es capaz de fijarse sólo en las células del sistema nervioso central y las vías digestivas de los primates. Se cree que la fijación en receptores refleja homologías configurativas fortuitas entre la estructura de la superficie del virión y un componente de la superficie celular. Por ejemplo, el virus de la inmunodeficiencia humana (HIV) se une al receptor CD4 en las células del sistema inmunitario y se cree que el virus de la rabia interactúa con los receptores de acetilcolina y que el virus de Epstein-Barr reconoce al receptor para el tercer componente del complemento en las células B. Es probable que cada célula susceptible posea por lo menos 100,000 sitios receptores para un virus dado.
ResponderEliminarDespués de la fijación, la partícula viral entra en la célula. Esta etapa se conoce como penetración, viropexia o fagocitosis. En algunos sistemas, se realiza por endocitosis mediada por receptores, con captación de las partículas virales ingeridas dentro de endosomas. Están menos claros los detalles de la penetración en otros sistemas. Ocurre perdida de la cubierta de manera concomitante con la penetración o poco después de la misma. La pérdida de la cubierta, o descubrimiento, es la separación física del ácido nucleico viral (o, en algunos casos, de las nucleocápsides inteARNs) a partir de los componentes estructurales externos del virión. En este momento se pierde la infecciosidad del virus original.
Los virus son los únicos agentes infecciosos en los que la disolución del agente infectante es una etapa obligatoria de la vía de la replicación
ResponderEliminarPoco después de quedar descubierto el genoma viral, se inicia la fase sintética del ciclo de replicación del virus. El aspecto esencial de la replicación viral consiste en que deben transcribirse los ARNm específicos del ácido nucleico viral para que se exprese y duplique con éxito la información genética. Una vez que se logra esta etapa, los virus recurren a los componentes de la célula para traducir el ARNm. Diversas clases de virus emplean vías diferentes para sintetizar los ARNm según la estructura del ácido nucleico viral. Algunos virus (por ejemplo, rhabdovirus, ortomixovirus y paramixovirus) poseen ARN polimerasas que sintetizan a los ARNm. Los virus ARN de este tipo se llaman virus de tira negativa (de sentido negativo), puesto que su genoma de ARN de una sola tira es complementario del ARNm, el cual se designa por acuerdo general como de tira positiva (o de sentido positivo).
Durante la replicación viral se sintetizan todas las macromoléculas especificadas del virus en una secuencia muy organizada. En algunas infecciones por virus, notablemente las producidas por virus de doble tira que contienen ADN, se sintetizan proteínas virales tempranas poco después de la infección y aparecen proteínas tardías sólo durante la fase tardía del proceso infeccioso, una vez que ha ocurrido la síntesis del ADN viral. Pueden activarse o no genes tempranos cuando se elaboran los productos tardíos. En contraste, se expresa al mismo tiempo la mayor parte de la información genética de los virus que contienen ARN o toda ella. Además de estos controles temporales, existen también controles cuantitativos puesto que no se elaboran todas las proteínas virales en las mismas cantidades. Las proteínas específicas del virus pueden regular la extensión de la transcripción del genoma o la traducción del ARNm viral.
Se encuentra la variación más amplia en las estrategias de la expresión genética entre los virus que contienen ARN. Algunos viriones poseen polimerasas (ortomixovirus, reovirus); otros sistemas emplean mensajes subgenómicos generados a veces por empalme (ortomixovirus, retrovirus); por último, algunos virus sintetizan grandes precursores poliproteínicos que se someten a procesamiento y segmentación para generar los productos génicos finales (picornavirus, retrovirus).
ResponderEliminarLa extensión a la que las enzimas específicas del virus participan en estos procesos varía de un grupo a otro. En general, los virus de mayor tamaño (herpesvirus, poxvirus) son más independientes de las funciones celulares que los virus de menor tamaño. Esta es una razón de que los virus de mayor tamaño sean más sensibles a la quimioterapia antiviral, porque se dispone de más procesos específicos de los virus como blancos de la acción farmacológica.
Los sitios intracelulares en los que ocurren los diferentes acontecimientos de la replicación viral, varían de un grupo a otro. Es posible plantear algunas generalizaciones. La proteína viral se sintetiza en el citoplasma, sobre los polirribosomas compuestos por ARNm específico del virus y ribosomas de la célula huésped. El ADN viral suele replicarse en el núcleo. El ARN genómico viral se duplica generalmente en el citoplasma celular, aunque hay excepciones.
#2012-1270 Etapas de la Replicación Viral
ResponderEliminarEl ciclo de multiplicación viral se define como la serie de etapas secuenciales que transcurren desde el contacto inicial de un virus con una célula huésped hasta la liberación a partir de la misma célula infectada, de la progenie de nuevos viriones.
El primer paso en la infección de una célula es la adhesión a la superficie celular. Se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos en el exterior de la célula. Las células que carecen de los receptores apropiados no son susceptibles al virus.
Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus.
(A) Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular.
(B) Entada vía endosomas en la superficie celular
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
3. PÉRDIDA DE LA CÁPSULA
El ácido nucleico debe de estar lo suficientemente expuesto como para que la replicación viral comience en esta fase. Cuando el ácido nucleico es expuesto, las partículas víricas infecciosas no pueden removerse de las células – este es el comienzo de la fase de ECLIPSE – la cual perdura hasta que viriones infecciosos nuevos sean creados.
4. SÍNTESIS DE ÁCIDO NUCLEICO Y PROTEINAS VIRALES
Existen diversos mecanismos, algunos serán abarcados en capítulos posteriores.
5. ENSAMBLAJE/MADURACIÓN
Nuevas partículas víricas son ensambladas. Puede haber un estadio de maduración posterior al proceso inicial de ensamblaje.
6. LIBERACIÓN O DESCARGA
Los virus pueden ser liberados mediante lisis celular, o, si son envueltos, pueden yemar de las células. Los virus que yeman no necesariamente destruyen la célula. Por tanto, algunos de estos virus son capaces de instaurar infecciones persistentes. No todas las partículas víricas liberadas son infecciosas. La proporción partículas infecciosas:partículas no-infecciosas varía según el virus y las condiciones de crecimiento.
#2012-1270 Adhesión
ResponderEliminarEl evento inicial en el ciclo de vida de un virus es la unión del mismo con la célula huésped a través del proceso de adsorción. Esta unión específica se establece entre una molécula presente en la membrana celular (receptor) y una proteína externa del virión. Algunos receptores virales han sido identificados y son en general carbohidratos, lípidos o proteínas de la membrana plasmática.
Así, la susceptibilidad o resistencia de una célula a la infección está determinada por la presencia o ausencia de receptores específicos. La importancia de la carencia de receptores se demuestra indirectamente porque es posible infectar exitosamente células no susceptibles a algunos virus utilizando ácido nucleico desnudo (transfección).
La unión virus-célula ocurre en dos etapas: una primera unión reversible por atracción electrostática, facilitada por la presencia de cationes: la adsorción irreversible se produce cuando se establece la unión polivalente virus-célula, ya que suele haber muchas copias de los receptores en la célula y múltiple sitios de unión en el virión.
#2012-1270 Penetración
ResponderEliminarUna vez establecida la adsorción, los virus pueden entrar en la célula a través de tres mecanismos diferentes: endocitosis mediada por receptor, fusión con la membrana plasmática y traslocación a través de la membrana plasmática.
Se acepta en la actualidad que los virus desnudos pueden entrar en la célula por traslocación o por endocitosis mientras que los virus envueltos penetran por endocitosis o por fusión.
En la endocitosis mediada por receptor el virus utiliza la misma vía de entrada que emplean muchas macromoléculas fisiológicamente importantes como nutrientes hormonas y factores de crecimiento. El virus absorbido a su receptor va quedando encerrado dentro de invaginaciones de la membrana plasmática recubiertas de una proteína llamada clatrina, que forman primero un hueco recubierto y luego una vesícula. De este modo el virión intacto entra en el endosoma. El pH ácido del endosoma (5 a 5,5) produce cambios conformacionales en las proteínas del virión que permiten la fusión de la membrana del endosoma con la membrana externa del virus y expulsan el genoma viral al citoplasma celular para su libre expresión.
El mecanismo de fusión directa de la envoltura viral con la membrana plasmática celular es utilizado por ciertos virus envueltos como herpes y Paramixovirus. Estos virus poseen en su envoltura glicoproteínas con capacidad de fusión independiente del pH, es decir, que a un pH neutro ocurre la fusión a nivel de la membrana celular liberando el genoma al citoplasma. A diferencia de la entrada por endocitosis, en este caso en ningún momento se ven viriones intactos dentro de la célula. Esta capacidad de fusión con membrana plasmática que tienen ciertos virus ha permitido su utilización como agentes fusogénicos para la obtención de híbridos celulares. (Ver virus del VIH)
Por último, algunos virus desnudos como Adenovirus parecen entrar directamente a través de la membrana plasmática mediante un proceso de traslocación.
#2012-1270 Penetración de los bacteriófagos
ResponderEliminarEl ciclo de replicación de un bacteriófago se puede dividir esquemáticamente en distintas etapas, las que son comunes a otros virus bacterianos y eucarióticos.
Adsorción: El virus se fija o adsorbe a componentes de la superficie celular que actúan como receptores específicos. La zona de adsorción del virus es complementaria al receptor celular, por lo tanto un determinado virus sólo puede infectar un número limitado de cepas celulares que contengan a un determinado receptor. La naturaleza de la zona de adsorción varía con el tipo de fago. En el T4 se localiza en el extremo de la cola, en donde se encuentran la placa basal, las espículas y las fibras de la cola.
Inyección del material genético viral: Después de la adsorción, se produce un cambio configuracional en las proteínas de la placa basal. Mediante su actividad enzimática producen un poro en la membrana citoplasmática de la célula. La vaina del fago se contrae y el material genético viral ingresa en la célula, mientras que la envoltura proteica queda en el exterior.
Replicación del material genético viral: El material genético viral que ingresa en una célula contiene bases modificadas que evitan la degradación por nucleasas bacterianas. Esta modificación consiste en la glicosilación y/o metilación de algunas determinadas bases. En el caso del fago T4 se glucosila la base 5'-hidroximetilcitosina. Se deben sintetizar proteínas tempranas que reparan el poro de la membrana citoplasmática por donde ingresó el genoma viral, degradan el DNA bacteriano lo que proporciona una fuente de precursores, evita la síntesis de RNA y proteínas bacterianas, y proporciona ribosomas para la síntesis de proteínas del fago. Además algunas de estas proteínas tempranas participan en la síntesis de las bases inusuales. La forma de replicación del genoma viral es dependiente del tipo de material genético (si es RNA o DNA, si es simple o doble cadena).
Síntesis de las envolturas proteicas: Las proteínas de la envoltura son proteínas tardías que se sintetizan después de iniciada la replicación del material genético.
Ensamble: Todas las proteínas de la envoltura se ensamblan para formar una partícula viral madura capaz de infectar a otra célula cuando sea liberada.
Lisis celular y liberación de las partículas virales: La lisis celular se debe a la síntesis de proteínas tardías codificadas en el genoma del fago.
#2012-1270 Replicación y maduración
ResponderEliminarREPLICACION DE VIRUS DNA
Después de la adsorción, penetración y eliminación de la cubierta, el ácido nucleico viral es liberado y migra al núcleo de la célula hospedadora. Se transcribe la porción de DNA de los genes tempranos. Tiene lugar la traducción, siendo los productos de estos genes enzimas requeridos para la multiplicación del DNA vírico. En la mayoría de los virus DNA, la transcripción temprana se lleva a cabo con la transcriptasa (RNA polimerasa) del hospedador. Tiempo después comienza la transcripción y traducción de los genes tardíos. La síntesis de las proteínas de la cápsida ocurre en el citoplasma de la célula hospedadora. Las proteínas de la cápsida migran al núcleo donde tiene lugar la maduración: el DNA vírico y las proteínas de la cápsida se ensamblan para formar las partículas víricas completas.
REPLICACION DE VIRUS RNA
a) VIRUS DE ARN QUE NO POSEEN UNA FASE DE ADN
Los virus que se replican vía intermediarios de ARN necesitan una ARN – polimerasa dependiente de ARN para replicar su ARN, pero las células animales aparentemente no tienen esa enzima. Por tanto, este tipo de virus de ARN necesita codificar su ARN – polimerasa dependiente de ARN.
Ninguna proteína viral puede ser sintetizada hasta que el ARN mensajero del virus esté disponible.
i. Virus de ARN de cadena positiva
En estos virus, el ARN del virión tiene el mismo sentido que un ARNm y funciona como tal. Este ARNm puede ser traducido inmediatamente luego de la infección.
ii. Virus de ARN de cadena negativa
El ARN del virión tiene sentido negativo (complementario al ARNm). Se copia en un ARNm de sentido positivo complementario antes de que las proteínas puedan ser sintetizadas. Entonces, necesitan codificar para una ARN – polimerasa dependiente de ARN y empacarla en el virión para sintetizar ARNm cuando infectan.
iii. Virus de ARN de cadena doble
No puede funcionar como ARNm; necesitan también empacar una ARN – polimerasa para sintetizar su ARNm luego de producir infecciones en células huésped.
b) VIRUS DE ARN QUE COPIAN SU ARN EN ADN.
Son los retrovirus de ARN sentido positivo que no funciona como ARNm luego de la infección puesto que no es liberado de su cápside al citoplasma. Sirve de plantilla para la transcriptasa inversa y es copiado en ADN. La enzima no está disponible en la célula se codifica y empaca en los viriones.
#2012-1270 Replicación y maduración
ResponderEliminarREPLICACION DE VIRUS DNA
Después de la adsorción, penetración y eliminación de la cubierta, el ácido nucleico viral es liberado y migra al núcleo de la célula hospedadora. Se transcribe la porción de DNA de los genes tempranos. Tiene lugar la traducción, siendo los productos de estos genes enzimas requeridos para la multiplicación del DNA vírico. En la mayoría de los virus DNA, la transcripción temprana se lleva a cabo con la transcriptasa (RNA polimerasa) del hospedador. Tiempo después comienza la transcripción y traducción de los genes tardíos. La síntesis de las proteínas de la cápsida ocurre en el citoplasma de la célula hospedadora. Las proteínas de la cápsida migran al núcleo donde tiene lugar la maduración: el DNA vírico y las proteínas de la cápsida se ensamblan para formar las partículas víricas completas.
REPLICACION DE VIRUS RNA
a) VIRUS DE ARN QUE NO POSEEN UNA FASE DE ADN
Los virus que se replican vía intermediarios de ARN necesitan una ARN – polimerasa dependiente de ARN para replicar su ARN, pero las células animales aparentemente no tienen esa enzima. Por tanto, este tipo de virus de ARN necesita codificar su ARN – polimerasa dependiente de ARN.
Ninguna proteína viral puede ser sintetizada hasta que el ARN mensajero del virus esté disponible.
i. Virus de ARN de cadena positiva
En estos virus, el ARN del virión tiene el mismo sentido que un ARNm y funciona como tal. Este ARNm puede ser traducido inmediatamente luego de la infección.
ii. Virus de ARN de cadena negativa
El ARN del virión tiene sentido negativo (complementario al ARNm). Se copia en un ARNm de sentido positivo complementario antes de que las proteínas puedan ser sintetizadas. Entonces, necesitan codificar para una ARN – polimerasa dependiente de ARN y empacarla en el virión para sintetizar ARNm cuando infectan.
iii. Virus de ARN de cadena doble
No puede funcionar como ARNm; necesitan también empacar una ARN – polimerasa para sintetizar su ARNm luego de producir infecciones en células huésped.
b) VIRUS DE ARN QUE COPIAN SU ARN EN ADN.
Son los retrovirus de ARN sentido positivo que no funciona como ARNm luego de la infección puesto que no es liberado de su cápside al citoplasma. Sirve de plantilla para la transcriptasa inversa y es copiado en ADN. La enzima no está disponible en la célula se codifica y empaca en los viriones.
#2012-1270 Maduración y salida de virus desnudos y envueltos
ResponderEliminarEl evento inicial en el ciclo de vida de un virus es la unión del mismo con la célula huésped a través de un receptor, proceso de adsorción. En la endocitosis mediada por receptor, el virus va quedando encerrado dentro de invaginaciones de la membrana. De este modo el virión intacto entra en el endosoma. En cambio, el mecanismo de fusión directa de la envoltura viral con la membrana plasmática celular es utilizado por ciertos virus envueltos que poseen glicoproteínas con capacidad de fusión independiente del pH. El desnudamiento ocurre simultáneamente o después de la penetración y comprende la eliminación de todas o parte de las cubiertas proteicas que rodean al ácido nucleico viral, de manera de dejar expuesto dentro de la célula el genoma viral desnudo o asociado a alguna enzima para que pueda expresarse. Se sintetizan proteínas virales y se replica el ácido nucleico viral.
Virus con genoma de ADN: Los virus llevan a cabo la duplicación de su ADN por medio de mecanismos y herramientas similares a los empleados por la célula para replicar su material genético. La replicación del ADN viral es generalmente semiconservativa, es decir cada cadena del ADN parental se conserva y por apareamiento con las bases complementarias se duplica, produciéndose como resultado final dos moléculas de ADN, cada una de las cuales está formada por una cadena de ADN parental y una cadena sintetizada "de novo".
Virus con genoma de ARN: A diferencia de lo que sucede con el ADN, la replicación del ARN es un fenómeno único de los virus ya que la maquinaria genética de la célula se basa exclusivamente en el uso del ADN como templado (ADN-ADN o ADN-ARN). Por lo tanto, para la duplicación de su genoma, así como acontecía para la síntesis de sus ARNm, los virus ARN deben necesariamente aportar la enzima ARN polimerasa ARN dependiente.
En las etapas tardías del ciclo de multiplicación viral las moléculas de ácido nucleico y proteínas virales sintetizadas en la célula infectada deben ensamblarse para formar los nuevos viriones y salir al espacio extracelular para estar en condiciones de infectar nuevas células. Este proceso de maduración y ensamble, también denominado morfogénesis, comprende el armado de la cápside a partir de las unidades estructurales y luego la formación del virión completo. El mecanismo va a diferir en base a dos propiedades del virión: la estructura de la cápside y la presencia o no de envoltura.
#2012-1270 Salida de un virus envuelto
ResponderEliminarLos virus envueltos en el último paso al ensamblaje, el virion, se une con la membrana celular a la salida de la célula, mediante proteínas que se insertan en la superficie interna y externa de la membrana. El virion naciente brota a través de la membrana. El ensamblaje y maduración con la expulsión a través de la superficie celular proporcionan un mecanismo más eficaz de salida, ya que no depende de la desintegración de la célula infectada. A la inserción de glicoproteínas virales en la superficie celular se cambia la especifidad antigénica, con lo que la célula infectada se convierte en blanco para el aparato inmunológico del huésped.
Wladimir A. Rijo Inirio 89257
ResponderEliminarD# 1(Etapas de la replicacion Viral)
Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada.
Wladimir A. Rijo Inirio 89257
ResponderEliminarD# 2(Adhesion)
El primer paso en la infección de una célula es la adhesión o fijación a la superficie celular. Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula. Las células que carecen de los receptores apropiados no son susceptibles al virus.
Wladimir A. Rijo Inirio 89257
ResponderEliminarD# 3 (Penetracion)
Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus.
Virus envueltos
(A) Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular.
(B) Entada vía endosomas en la superficie celular.
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
Virus no envueltos o desnudos
Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
Wladimir A. Rijo Inirio 89257
ResponderEliminarD# 4(Penetracion de los Bacteriografos)
Los virus complejos producen una rotura en la membrana celular del hospedador en uno de los puntos de anclaje, gracias a la presencia de algunas moléculas de enzimas hidrolíticas entre las proteínas de la cápside. A través de la rotura, el tubo central inyecta el ADN vírico, quedando la cápside vacía en el exterior de la célula diana y el ácido nucleico libre en el citoplasma. La presencia de cápsidas en la superficie celular es un buen indicio de que ha sufrido una infección vírica.
Otros virus sin envoltura lipídica se introducen en la célula con cápsida y todo, lo cual puede realizarse de dos maneras:
Por penetración directa: después de la fijación, el virus abre una brecha en la membrana y se introduce en el citoplasma.
Por endocitosis: la membrana forma una invaginación en torno al virus, llegando a formar una vesícula que penetra en la célula. Formada la vesícula, el virus abre una brecha en la membrana de la misma con ayuda de algunas enzimas hidrolíticas que él mismo transporta, penetrando así en el citoplasma.
Los virus con envoltura lipídica burlan la barrera de la membrana celular porque su cubierta lipídica se funde con la membrana, ya que son de la misma naturaleza. Esta fusión de membranas puede realizarse en dos lugares distintos:
Fusión en la superficie celular: de manera que el virión penetra directamente en el citoplasma.
Fusión con un lisosoma: se forma una vesícula por endocitosis, a la que se une un lisosoma para digerir la partícula introducida; entonces, la cubierta lipídica del virus se funde con la membrana del lisosoma y el virión escapa hacia el citoplasma.
Wladimir A. Rijo Inirio 89257
ResponderEliminarD# 5 (Replicacion y Maduracion)
Ensamblaje de los componentes de los virus para formar nuevas partículas víricas (proceso también llamado maduración).
Los virus desnudos realizan su ensamblaje y maduración en forma intracelular y adquieren capacidad infectante dentro de la célula. Para su salida requieren de la desintegración celular. Los virus envueltos en el último paso al ensamblaje el virion se une con la membrana celular a la salida de la célula, mediante proteínas que se insertan en la superficie interna y exterior de la membrana. El virion naciente brota atraves de la membrana. El ensamblaje y maduración con la expulsión atraves de la superficie celular proporcionan un mecanismo más eficaz de salida, ya que no depende de la desintegración de la célula infectada.
Wladimir A. Rijo Inirio 89257
ResponderEliminarD# 6(Maduracion y salida de virus desnudos y envueltos)
Después de la multiplicación del virus tiene lugar la salida de los nuevos individuos, que saldrán con capacidad de infectar nuevas células. Las principales modalidades de liberación da nombre a nuevas variantes del ciclo vital de los virus:
Infección persistente
Los nuevos virus no esperan a la muerte de la célula hospedadora para abandonarla, sino que van saliendo de la célula al mismo tiempo que se van produciendo, de manera que la célula puede seguir viva y produciendo nuevas partículas víricas. La liberación puede hacerse de dos maneras:
Los virus sin envoltura lipoproteica salen directamente, sin arrastrar ningún resto de la membrana plasmática, bien sea abriendo una brecha en la membrana, o bien aprovechando los mecanismos de exocitosis o salida de sustancias al exterior de la célula.
Wladimir A. Rijo Inirio 89257
ResponderEliminarD# 7(Salida de virus envuelto)
Liberación de los nuevos virus de la célula hospedadora, que ya pueden infectar otras células. Pueden producirse con o sin rotura de la célula, dependiendo de los virus. Los virus desnudos salen por lisis celular lo contrario de los envueltos que salen por gemación.
88032
ResponderEliminarSlide # 2 y3 Adhesion y Penetracion
El primer paso en la infección de una célula es la adhesión o fijación a la superficie celular. Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula. Las células que carecen de los receptores apropiados no son susceptibles al virus.
La penetración: Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus.Virus envueltos Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular. Entada vía endosomas en la superficie celular. Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
Virus no envueltos o desnudos
Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
88032
ResponderEliminarslide #4
La primera etapa de la infección viral es la interacción de un virión con un sitio receptor específica de la superficie celular. Las moléculas receptoras difieren para los distintos virus, y son proteínas en algunos casos (por ejemplo, picornavirus) y oligosacáridos en otros (por ejemplo, ortovirus y paramixovirus). La presencia o la ausencia de receptores desempeña una función determinante de primer orden en el tropismo celular y la patogenia viral; por ejemplo, el poliovirus es capaz de fijarse sólo en las células del sistema nervioso central y las vías digestivas de los primates. Se cree que la fijación en receptores refleja homologías configurativas fortuitas entre la estructura de la superficie del virión y un componente de la superficie celular.
El virus de la inmunodeficiencia humana (HIV) se une al receptor CD4 en las células del sistema inmunitario y se cree que el virus de la rabia interactúa con los receptores de acetilcolina y que el virus de Epstein-Barr reconoce al receptor para el tercer componente del complemento en las células B. Es probable que cada célula susceptible posea por lo menos 100,000 sitios receptores para un virus dado
88032
ResponderEliminarSlide #5 Replicacion y Maduracion
Ensamblaje de los componentes de los virus para formar nuevas partículas víricas.
Los virus desnudos realizan su ensamblaje y maduración en forma intracelular y adquieren capacidad infectante dentro de la célula. Para su salida requieren de la desintegración celular. Los virus envueltos en el último paso al ensamblaje el virion se une con la membrana celular a la salida de la célula, mediante proteínas que se insertan en la superficie interna y exterior de la membrana. El virion naciente brota atraves de la membrana. El ensamblaje y maduración con la expulsión atraves de la superficie celular proporcionan un mecanismo más eficaz de salida, ya que no depende de la desintegración de la célula infectada.
88032
ResponderEliminarSlide #6 Maduracion y Lberacion
Los virus en general pueden liberarse de la célula mediante dos tipos de mecanismos: lisis y gemación.
Para los virus líticos -la mayoría de los virus desnudos-, la liberación es un proceso simple: la célula infectada se rompe y libera las partículas virales. En el caso de los virus que se liberan por gemación, estos adquieren su envoltura lipídica y las proteínas asociadas en el momento en que pasan a través de la membrana plasmática o de vesículas intracelulares. La liberación de los virus por esta vía puede resultar altamente perjudicial para la célula -por ej.en el caso de paramixovirus, rabdovirus, togavirus- o no -por ej. retrovirus-; de todos modos en cualquiera de los casos el proceso es controlado por el virus: la interacción física de las proteínas de la cápside sobre la superficie interna de la membrana celular fuerza a la partícula viral a pasar a través de dicha membrana.
El ensamble, la maduración y liberación son generalmente procesos simultáneos en los virus que se liberan por gemación. El tipo de membrana a través de la cual geman los virus depende del tipo de virus, por ej. puede ser a través de la membrana celular como en el caso de los retrovirus y togavirus, a través de la membrana citoplasmática -ortomixovirus, coronavirus, bunyavirus, rabdovirus, hepadnavirus- o a través de la membrana nuclear como en los herpesvirus.
Slide 7: Salida de un virus envuelto 2012-0593
Se encuentra la variación más amplia en las estrategias de la expresión genética entre los virus que contienen ARN. Algunos viriones poseen polimerasas (ortomixovirus, reovirus); otros sistemas emplean mensajes subgenómicos generados a veces por empalme (ortomixovirus, retrovirus); por último, algunos virus sintetizan grandes precursores poliproteínicos que se someten a procesamiento y segmentación para generar los productos génicos finales (picornavirus, retrovirus).Los sitios intracelulares en los que ocurren los diferentes acontecimientos de la replicación viral, varían de un grupo a otro. Es posible plantear algunas generalizaciones. La proteína viral se sintetiza en el citoplasma, sobre los polirribosomas compuestos por ARNm específico del virus y ribosomas de la célula huésped. El ADN viral suele replicarse en el núcleo.
88032
ResponderEliminarSlide #7 Salida del virus envuelto
Liberación de los nuevos virus de la célula hospedadora, que ya pueden infectar otras células. Pueden producirse con o sin rotura de la célula, dependiendo de los virus. Los virus desnudos salen por lisis celular lo contrario de los envueltos que salen por gemación.
-todo Se encuentra la variación más amplia en las estrategias de la expresión genética entre los virus que contienen ARN. Algunos viriones poseen polimerasas (ortomixovirus, reovirus); otros sistemas emplean mensajes subgenómicos generados a veces por empalme (ortomixovirus, retrovirus). Dependiendo del virus tiene distintos manera de salida virus desnudos salen por lisis celular y los envueltos que salen por gemación.
1.Etapas de la replicacion de virosis
ResponderEliminarEl Ciclo De Replicación De Los Virus consta generalmente de las siguientes fases: fijación y entrada en la célula, eclipse, multiplicación y liberación del virus.
2.Adhesion
ResponderEliminarEl primer paso en la infección viral es la absorción a la membrana de la célula susceptible por medio de la adhesión de ligandos virales (proteínas de la capside o glucoproteínas de las espículas, por ejemplo a receptores superficiales de la célula). La distribución en el cuerpo del hospedador de ciertos tipos de receptores celulares explica el tropismo tisular y de hospedador de los virus. Los tejidos y células que carecen de receptores específicos de virus determinados no son infectados por dichos viriones. La naturaleza de los receptores es variable y ciertos virus pueden tener más de un tipo de receptor.
3.Penetracion
ResponderEliminarLos virus complejos producen una rotura en la membrana celular del hospedador en uno de los puntos de anclaje, gracias a la presencia de algunas moléculas de enzimas hidrolíticas entre las proteínas de la cápsida. A través de la rotura, el tubo central inyecta el ADN vírico, quedando la cápside vacía en el exterior de la célula diana y el ácido nucleico libre en el citoplasma. La presencia de cápsidas en la superficie celular es un buen indicio de que ha sufrido una infección vírica. Otros virus sin envoltura lipídica se introducen en la célula con cápsida y todo, lo cual puede realizarse de dos maneras:
Por penetración directa: después de la fijación, el virus abre una brecha en la membrana y se introduce en el citoplasma.
Por endocitosis: la membrana forma una invaginación en torno al virus, llegando a formar una vesícula que penetra en la célula. Formada la vesícula, el virus abre una brecha en la membrana de la misma con ayuda de algunas enzimas hidrolíticas que él mismo transporta, penetrando así en el citoplasma.
4.Penetracion de los bacteriofagos
ResponderEliminarAdsorción: El virus se fija o adsorbe a componentes de la superficie celular que actúan como receptores específicos. La zona de adsorción del virus es complementaria al receptor celular, por lo tanto un determinado virus sólo puede infectar un número limitado de cepas celulares que contengan a un determinado receptor. La naturaleza de la zona de adsorción varía con el tipo de fago. En el T4 se localiza en el extremo de la cola, en donde se encuentran la placa basal, las espículas y las fibras de la cola.
Inyección del material genético viral: Después de la adsorción, se produce un cambio configuracional en las proteínas de la placa basal, alguna de las cuales tienen actividad enzimática y producen un poro en la membrana citoplasmática de la célula. La vaina del fago se contrae y el material genético viral ingresa en la célula, mientras que la envoltura proteica queda en el exterior.
Replicación del material genético viral: El material genético viral que ingresa en una célula contiene bases modificadas que evitan la degradación por nucleasas bacterianas. Esta modificación consiste en la glicosilación y/o metilación de algunas determinadas bases. En el caso del fago T4 se glucosila la base 5'-hidroximetilcitosina. Para lograr una efectiva replicación del genoma viral se deben sintetizar algunas proteínas ni bien el material genético ingresa en la célula.
Síntesis de las envolturas proteicas: Las proteínas de la envoltura (cápside, vaina, fibras, etc) son proteínas tardías que se sintetizan después de iniciada la replicación del material genético. La síntesis de cada componente proteico se realiza separadamente. En el caso del fago T4, el material genético es encapsidado antes del ensamble del resto de los componentes.
Ensamble: Todas las proteínas de la envoltura se ensamblan para formar una partícula viral madura capaz de infectar a otra célula cuando sea liberada.
Lisis celular y liberación de las partículas virales: La lisis celular se debe a la síntesis de proteínas tardías codificadas en el genoma del fago. En el fago T4, estas proteínas son enzimas que lesionan la membrana citoplasmática y la pared celular.
5.Replicacion y Maduracion
ResponderEliminarLa multiplicación del virus consta de la replicación de su material genético, de la transcripción de su mensaje en una molécula de ARN y de la traducción del mensaje para producir proteínas víricas, tanto las que formarán parte de la cápsida como las proteínas enzimáticas necesarias para el ensamblaje de las piezas del virión y para algunas de las funciones anteriores. Los ribosomas y la mayor parte de las enzimas que los ácidos nucleicos víricos utilizan en estos procesos son los de la célula infectada. Posteriormente a estos procesos, tiene lugar el ensamblaje de las piezas para construir nuevos viriones. En muchos virus, como el del mosaico del tabaco, el ensamblaje es automático y depende de la concentración salina del medio. En otros virus, en el ensamblaje intervienen enzimas codificadas en el ácido nucleico del virus.
6.Maduracion y salida de virus envueltos y desnudos
ResponderEliminarLos virus desnudos realizan su ensamble y maduracion en forma intracelular, y adquieren capacidad infectante dentro de la celula. Para su salida requieren de la desintegracion celular. La mayoría de los virus desnudos son resistentes al medio externo, a la desecación y a los solventes de lípidos. Por esta razón, los virus desnudos pueden penetrar por vía digestiva ya que resisten el pH ácido del estómago. Pueden ser transmitidos por vía fecal.oral, a través de agua o alimentos contaminados o por manos u objetos. Son ejemplos de virus desnudos el Poliovirus, y los Adenovirus. Los virus envueltos en el ultimo paso al ensamble, se unen con la membrana celular a la salida de la celula, mediante proteinas que se insertan en la superficie interna y exterior de la membrana.
7.Salida de un virus envuelto
ResponderEliminarLos virus envueltos en el ultimo paso al ensamble (el virion), se unen con la membrana celular a la salida de la celula, mediante proteinas que se insertan en la superficie interna y exterior de la membrana. Al final se obtiene un virion infeccioso con envoltura de glicoproteinas.
87451 Felix Bienvenido Carrasco Vasquez grupo: lunes 10-12
ResponderEliminarAdhesión
La adhesión es la propiedad de la materia por la cual se unen y plasman dos superficies de sustancias iguales o diferentes cuando entran en contacto, y se mantienen juntas por fuerzas intermoleculares.
La adhesión ha jugado un papel muy importante en muchos aspectos de las técnicas de construcción tradicionales. La adhesión del ladrillo con el mortero (cemento) es un ejemplo claro.
La cohesión es distinta de la adhesión. La cohesión es la fuerza de atracción entre partículas adyacentes dentro de un mismo cuerpo, mientras que la adhesión es la interacción entre las superficies de distintos cuerpos.
87451 Felix Bienvenido Carrasco Vasquez grupo: lunes 10-12
ResponderEliminarPenetración
Los virus complejos producen una rotura en la membrana celular del hospedador en uno de los puntos de anclaje, gracias a la presencia de algunas moléculas de enzimas hidrolíticas entre las proteínas de la cápsida. A través de la rotura, el tubo central inyecta el ADN vírico, quedando la cápside vacía en el exterior de la célula diana y el ácido nucleico libre en el citoplasma. La presencia de cápsidas en la superficie celular es un buen indicio de que ha sufrido una infección vírica. Otros virus sin envoltura lipídica se introducen en la célula con cápsida y todo, lo cual puede realizarse de dos maneras:
Por penetración directa: después de la fijación, el virus abre una brecha en la membrana y se introduce en el citoplasma.
Por endocitosis: la membrana forma una invaginación en torno al virus, llegando a formar una vesícula que penetra en la célula. Formada la vesícula, el virus abre una brecha en la membrana de la misma con ayuda de algunas enzimas hidrolíticas que él mismo transporta, penetrando así en el citoplasma.
87451 Felix Bienvenido Carrasco Vasquez grupo: lunes 10-12
ResponderEliminarPenetracion de los bacteriofagos
Adsorción: El virus se fija o adsorbe a componentes de la superficie celular que actúan como receptores específicos. La zona de adsorción del virus es complementaria al receptor celular, por lo tanto un determinado virus sólo puede infectar un número limitado de cepas celulares que contengan a un determinado receptor. La naturaleza de la zona de adsorción varía con el tipo de fago. En el T4 se localiza en el extremo de la cola, en donde se encuentran la placa basal, las espículas y las fibras de la cola.
Inyección del material genético viral: Después de la adsorción, se produce un cambio configuracional en las proteínas de la placa basal, alguna de las cuales tienen actividad enzimática y producen un poro en la membrana citoplasmática de la célula. La vaina del fago se contrae y el material genético viral ingresa en la célula, mientras que la envoltura proteica queda en el exterior.
Replicación del material genético viral: El material genético viral que ingresa en una célula contiene bases modificadas que evitan la degradación por nucleasas bacterianas. Esta modificación consiste en la glicosilación y/o metilación de algunas determinadas bases. En el caso del fago T4 se glucosila la base 5'-
hidroximetilcitosina. Para lograr una efectiva replicación del genoma viral se deben sintetizar algunas proteínas ni bien el material genético ingresa en la célula.
Síntesis de las envolturas proteicas: Las proteínas de la envoltura (cápside, vaina, fibras, etc) son proteínas tardías que se sintetizan después de iniciada la replicación del material genético. La síntesis de cada componente proteico se realiza separadamente. En el caso del fago T4, el material genético es encapsidado antes del ensamble del resto de los componentes.
Ensamble: Todas las proteínas de la envoltura se ensamblan para formar una partícula viral madura capaz de infectar a otra célula cuando sea liberada.
Lisis celular y liberación de las partículas virales: La lisis celular se debe a la síntesis de proteínas tardías codificadas en el genoma del fago. En el fago T4, estas proteínas son enzimas que lesionan la membrana citoplasmática y la pared celular.
87451 Felix Bienvenido Carrasco Vasquez grupo: lunes 10-12
ResponderEliminarReplicacion y Maduracion
La multiplicación del virus consta de la replicación de su material genético, de la transcripción de su mensaje en una molécula de ARN y de la traducción del mensaje para producir proteínas víricas, tanto las que formarán parte de la cápsida como las proteínas enzimáticas necesarias para el ensamblaje de las piezas del virión y para algunas de las funciones anteriores. Los ribosomas y la mayor parte de las enzimas que los ácidos nucleicos víricos utilizan en estos procesos son los de la célula infectada. Posteriormente a estos procesos, tiene lugar el ensamblaje de las piezas para construir nuevos viriones. En muchos virus, como el del mosaico del tabaco, el ensamblaje es automático y depende de la concentración salina del medio. En otros virus, en el ensamblaje intervienen enzimas codificadas en el ácido nucleico del virus.
87451 Felix Bienvenido Carrasco Vasquez grupo: lunes 10-12
ResponderEliminarMaduracion y salida de virus envueltos y desnudos
Los virus desnudos realizan su ensamble y maduracion en forma intracelular, y adquieren capacidad infectante dentro de la celula. Para su salida requieren de la desintegracion celular. La mayoría de los virus desnudos son resistentes al medio externo, a la desecación y a los solventes de lípidos. Por esta razón, los virus desnudos pueden penetrar por vía digestiva ya que resisten el pH ácido del estómago. Pueden ser transmitidos por vía fecal.oral, a través de agua o alimentos contaminados o por manos u objetos. Son ejemplos de virus desnudos el Poliovirus, y los Adenovirus. Los virus envueltos en el ultimo paso al ensamble, se unen con la membrana celular a la salida de la celula, mediante proteinas que se insertan en la superficie interna y exterior de la membrana.
89348
ResponderEliminarETAPAS DE LA REPLICACIÓN VIRAL
1- El primer paso en la infección viral es la absorción a la membrana de la célula susceptible por medio de la adhesión de ligandos virales (proteínas de la capside o glucoproteínas de las espículas, por ejemplo a receptores superficiales de la célula). La distribución en el cuerpo del hospedador de ciertos tipos de receptores celulares explica el tropismo tisular y de hospedador de los virus. Los tejidos y células que carecen de receptores específicos de virus determinados no son infectados por dichos viriones. La naturaleza de los receptores es variable y ciertos virus pueden tener más de un tipo de receptor.
2- Los virus complejos producen una rotura en la membrana celular del hospedador en uno de los puntos de anclaje, gracias a la presencia de algunas moléculas de enzimas hidrolíticas entre lasproteínas de la cápsida. A través de la rotura, el tubo central inyecta el ADN vírico, quedando la cápside vacía en el exterior de la célula diana y el ácido nucleico libre en el citoplasma. La presencia de cápsidas en la superficie celular es un buen indicio de que ha sufrido una infección vírica.
Otros virus sin envoltura lipídica se introducen en la célula con cápsida y todo, lo cual puede realizarse de dos maneras:
• Por penetración directa: después de la fijación, el virus abre una brecha en la membrana y se introduce en el citoplasma.
• Por endocitosis: la membrana forma una invaginación en torno al virus, llegando a formar una vesícula que penetra en la célula. Formada la vesícula, el virus abre una brecha en la membrana de la misma con ayuda de algunas enzimas hidrolíticas que él mismo transporta, penetrando así en el citoplasma.
Los virus con envoltura lipídica burlan la barrera de la membrana celular porque su cubierta lipídica se funde con la membrana, ya que son de la misma naturaleza. Esta fusión de membranas puede realizarse en dos lugares distintos:
• Fusión en la superficie celular: de manera que el virión penetra directamente en el citoplasma.
• Fusión con un lisosoma: se forma una vesícula por endocitosis, a la que se une un lisosoma para digerir la partícula introducida; entonces, la cubierta lipídica del virus se funde con la membrana del lisosoma y el virión escapa hacia el citoplasma.
3- Según la duración de la fase de eclipse, se suelen distinguir dos modalidades de ciclo infeccioso de un virus:
• Ciclo ordinario: el ácido nucleico vírico procede inmediatamente a la transcripción de su mensaje genético en los ARN necesarios para su multiplicación, y prosigue rápidamente el ciclo vital. Este tipo de ciclo es el más extendido en la naturaleza.
• Ciclo lisogénico: fue descubierto por André Lwoff en bacteriófagos. El ADN vírico se cierra por sus extremos generando un ADN circular. Este ADN se inserta en el ADN bacteriano en un lugar específico en el que la secuencia de nucleótidos bacterianos es semejante alguna región del ADN vírico.
Algunos virus que infectan células animales siguen también el ciclo lisogénico, como los papilomavirus de las verrugas y algunos retrovirus que producen algunos tipos de cáncer.
En el caso de los retrovirus, conviene recordar que el ácido nucleico es ARN monocatenario, por lo que la transcriptasa inversa ha de copiar el genoma vírico en forma de ADN antes de que pueda insertarse en el ADN celular.
4- La multiplicación del virus consta de la replicación de su material genético, de la transcripción de su mensaje en una molécula de ARN y de la traducción del mensaje para producir proteínasvíricas, tanto las que formarán parte de la cápsida como las proteínas enzimáticas necesarias para el ensamblaje de las piezas del virión y para algunas de las funciones anteriores. Losribosomas y la mayor parte de las enzimas que los ácidos nucleicos víricos utilizan en estos procesos son los de la célula infectada.
2012-0276
ResponderEliminarLos virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada.
La única característica de la multiplicación viral consiste en que, poco después de su interacción con una célula huésped, el virión infectante se desintegra y pierde la infecciosidad que se puede medir. Esta fase del ciclo celular se llama periodo de eclipse; su duración varía dependiendo del virus y de la célula huésped en particular, y termina con la formación de las primeras partículas virales descendientes infecciosas. El periodo de eclipse es, en realidad, de intensa actividad sintética, puesto que la célula se redirige hacia la satisfacción de las necesidades del "pirata" viral. En algunos casos, tan pronto como el ácido nucleico viral penetra a la célula huésped, el metabolismo celular es recanalizado exclusivamente hacia la síntesis de nuevas partículas virales. En otros casos, los procesos metabólicos de la célula huésped no se alteran significativamente, aunque la célula sintetiza proteínas virales y ácidos nucleicos.
Los virus han desarrollado diversas estrategias para lograr la multiplicación en las células huéspedes parasitadas por ellos. Aunque los detalles varían de un grupo a otro, el patrón general de los ciclos de replicación es semejante. La primera etapa de la infección viral es la interacción de un virión con un sitio receptor específica de la superficie celular. Las moléculas receptoras difieren para los distintos virus, y son proteínas en algunos casos (por ejemplo, picornavirus) y oligosacáridos en otros (por ejemplo, ortovirus y paramixovirus). La presencia o la ausencia de receptores desempeña una función determinante de primer orden en el tropismo celular y la patogenia viral; por ejemplo, el poliovirus es capaz de fijarse sólo en las células del sistema nervioso central y las vías digestivas de los primates. Se cree que la fijación en receptores refleja homologías configurativas fortuitas entre la estructura de la superficie del virión y un componente de la superficie celular. Por ejemplo, el virus de la inmunodeficiencia humana (HIV) se une al receptor CD4 en las células del sistema inmunitario y se cree que el virus de la rabia interactúa con los receptores de acetilcolina y que el virus de Epstein-Barr reconoce al receptor para el tercer componente del complemento en las células B. Es probable que cada célula susceptible posea por lo menos 100,000 sitios receptores para un virus dado.
2012-0276
ResponderEliminarDespués de la fijación, la partícula viral entra en la célula. Esta etapa se conoce como penetración, viropexia o fagocitosis. En algunos sistemas, se realiza por endocitosis mediada por receptores, con captación de las partículas virales ingeridas dentro de endosomas. Están menos claros los detalles de la penetración en otros sistemas. Ocurre perdida de la cubierta de manera concomitante con la penetración o poco después de la misma. La pérdida de la cubierta, o descubrimiento, es la separación física del ácido nucleico viral (o, en algunos casos, de las nucleocápsides inteARNs) a partir de los componentes estructurales externos del virión. En este momento se pierde la infecciosidad del virus original. Los virus son los únicos agentes infecciosos en los que la disolución del agente infectante es una etapa obligatoria de la vía de la replicación. Poco después de quedar descubierto el genoma viral, se inicia la fase sintética del ciclo de replicación del virus. El aspecto esencial de la replicación viral consiste en que deben transcribirse los ARNm específicos del ácido nucleico viral para que se exprese y duplique con éxito la información genética. Una vez que se logra esta etapa, los virus recurren a los componentes de la célula para traducir el ARNm. Diversas clases de virus emplean vías diferentes para sintetizar los ARNm según la estructura del ácido nucleico viral. Algunos virus (por ejemplo, rhabdovirus, ortomixovirus y paramixovirus) poseen ARN polimerasas que sintetizan a los ARNm. Los virus ARN de este tipo se llaman virus de tira negativa (de sentido negativo), puesto que su genoma de ARN de una sola tira es complementario del ARNm, el cual se designa por acuerdo general como de tira positiva (o de sentido positivo). Durante la replicación viral se sintetizan todas las macromoléculas especificadas del virus en una secuencia muy organizada. En algunas infecciones por virus, notablemente las producidas por virus de doble tira que contienen ADN, se sintetizan proteínas virales tempranas poco después de la infección y aparecen proteínas tardías sólo durante la fase tardía del proceso infeccioso, una vez que ha ocurrido la síntesis del ADN viral. Pueden activarse o no genes tempranos cuando se elaboran los productos tardíos. En contraste, se expresa al mismo tiempo la mayor parte de la información genética de los virus que contienen ARN o toda ella. Además de estos controles temporales, existen también controles cuantitativos puesto que no se elaboran todas las proteínas virales en las mismas cantidades. Las proteínas específicas del virus pueden regular la extensión de la transcripción del genoma o la traducción del ARNm viral.
2012-0276
ResponderEliminarSe encuentra la variación más amplia en las estrategias de la expresión genética entre los virus que contienen ARN. Algunos viriones poseen polimerasas (ortomixovirus, reovirus); otros sistemas emplean mensajes subgenómicos generados a veces por empalme (ortomixovirus, retrovirus); por último, algunos virus sintetizan grandes precursores poliproteínicos que se someten a procesamiento y segmentación para generar los productos génicos finales (picornavirus, retrovirus). La extensión a la que las enzimas específicas del virus participan en estos procesos varía de un grupo a otro. En general, los virus de mayor tamaño (herpesvirus, poxvirus) son más independientes de las funciones celulares que los virus de menor tamaño. Esta es una razón de que los virus de mayor tamaño sean más sensibles a la quimioterapia antiviral, porque se dispone de más procesos específicos de los virus como blancos de la acción farmacológica. Los sitios intracelulares en los que ocurren los diferentes acontecimientos de la replicación viral, varían de un grupo a otro. Es posible plantear algunas generalizaciones. La proteína viral se sintetiza en el citoplasma, sobre los polirribosomas compuestos por ARNm específico del virus y ribosomas de la célula huésped. El ADN viral suele replicarse en el núcleo. El ARN genómico viral se duplica generalmente en el citoplasma celular, aunque hay excepciones.
89316
ResponderEliminarReplicación y maduración
Poco después de quedar descubierto el genoma viral, se inicia la fase sintética del ciclo de replicación del virus. El aspecto esencial de la replicación viral consiste en que deben transcribirse los ARNm específicos del ácido nucleico viral para que se exprese y duplique con éxito la información genética. Una vez que se logra esta etapa, los virus recurren a los componentes de la célula para traducir el ARNm. Diversas clases de virus emplean vías diferentes para sintetizar los ARNm según la estructura del ácido nucleico viral. Algunos virus (por ejemplo, rhabdovirus, ortomixovirus y paramixovirus) poseen ARN polimerasas que sintetizan a los ARNm. Los virus ARN de este tipo se llaman virus de tira negativa (de sentido negativo), puesto que su genoma de ARN de una sola tira es complementario del ARNm, el cual se designa por acuerdo general como de tira positiva (o de sentido positivo).
Durante la replicación viral se sintetizan todas las macromoléculas especificadas del virus en una secuencia muy organizada. En algunas infecciones por virus, notablemente las producidas por virus de doble tira que contienen ADN, se sintetizan proteínas virales tempranas poco después de la infección y aparecen proteínas tardías sólo durante la fase tardía del proceso infeccioso, una vez que ha ocurrido la síntesis del ADN viral. Pueden activarse o no genes tempranos cuando se elaboran los productos tardíos. En contraste, se expresa al mismo tiempo la mayor parte de la información genética de los virus que contienen ARN o toda ella. Además de estos controles temporales, existen también controles cuantitativos puesto que no se elaboran todas las proteínas virales en las mismas cantidades. Las proteínas específicas del virus pueden regular la extensión de la transcripción del genoma o la traducción del ARNm viral.
89316
ResponderEliminarBuenos modelos para el estudio de la expresión genética son los virus y los bacteriófagos pequeños de los animales. Su pequeño tamaño ha permitido aclarar la secuencia total de nucleótidos de unos cuantos virus. Esto ha hecho que se descubran genes sobrepuestos en los que se emplean algunas secuencias de ADN para la síntesis de dos poli péptidos diferentes, ya sea mediante el empleo de dos armazones de lectura diferentes o por medio de dos moléculas de ARNm que emplean los mismos armazones de lectura pero puntos de iniciación diferentes. Un sistema viral (adenovirus) reveló por primera vez el fenómeno de procesamiento de ARNm que se llama "empalme", por medio del cual se generan las secuencias de ARNm que codifican una proteína determinada a partir de secuencias separadas del modelo, con empalme de las secuencias no codificadoras intermedias de la transcripción.
Se encuentra la variación más amplia en las estrategias de la expresión genética entre los virus que contienen ARN. Algunos viriones poseen polimerasas (ortomixovirus, reovirus); otros sistemas emplean mensajes subgenómicos generados a veces por empalme (ortomixovirus, retrovirus); por último, algunos virus sintetizan grandes precursores poliproteínicos que se someten a procesamiento y segmentación para generar los productos génicos finales (picornavirus, retrovirus).
La extensión a la que las enzimas específicas del virus participan en estos procesos varía de un grupo a otro. En general, los virus de mayor tamaño (herpesvirus, poxvirus) son más independientes de las funciones celulares que los virus de menor tamaño. Esta es una razón de que los virus de mayor tamaño sean más sensibles a la quimioterapia antiviral, porque se dispone de más procesos específicos de los virus como blancos de la acción farmacológica.
Los sitios intracelulares en los que ocurren los diferentes acontecimientos de la replicación viral, varían de un grupo a otro. Es posible plantear algunas generalizaciones. La proteína viral se sintetiza en el citoplasma, sobre los poli ribosomas compuestos por ARNm específico del virus y ribosomas de la célula huésped. El ADN viral suele replicarse en el núcleo. El ARN genómico viral se duplica generalmente en el citoplasma celular, aunque hay excepciones.
89316
ResponderEliminarMaduración y salida de virus desnudos y envueltos
En las etapas tardías del ciclo de multiplicación viral las moléculas de ácido nucleico y proteínas virales sintetizadas en la célula infectada deben ensamblarse para formar los nuevos viriones y salir al espacio extracelular para estar en condiciones de infectar nuevas células. Este proceso de maduración y ensamble, también denominado morfogénesis, comprende el armado de la cápside a partir de las unidades estructurales y luego la formación del virión completo. El mecanismo va a diferir en base a dos propiedades del virión: la estructura de la cápside y la presencia o no de envoltura.
La maduración: El mecanismo mediante el cual las cápsidas se ensamblan alrededor de losácidos nucleicos no se conoce tan bien como la maduración de los fagos. Parece ser un proceso espontáneo en el que podrían estar implicadas algunas proteínas de la célula hospedadora.
La liberación: Los virus envueltos adquieran su cubierta en el proceso de salida de la célula hospedadora. Antes de que el virus deje la célula, los genes víricos hacen que la célula hospedadora sintetice las proteínas quedan embebidas en la región de la membrana plasmática de la célula, que constituirá la cubierta vírica. El virus se sitúa debajo de dicha región de la membrana plasmática de la célula, que constituirá la cubierta vírica. El virus se sitúa debajo de dicha región de la membrana; la empuja hacia fuera formando una yema o protuberancia y finalmente, sale de la célula mediante un proceso de exocitosis. Como consecuencia de esta salida, el virión de un virus envuelto es una estructura compuesta; es decir, el contenido del virión y la mayor parte de las proteínas de su membrana son codificados por el genoma vírico, pero la porción lípida está codificada por el genoma de la célula hospedadora. Los virus desnudos se liberan por un mecanismo más simple, similar al que realiza el fago T4.Finalizad el ensamblaje, los viriones se acumulan en el citoplasma, la célula se isa y muere liberando los viriones.
89316
ResponderEliminarSalida de un virus envuelto
Los genes víricos no se expresan hasta que el virus pierde sus cubiertas. Las membranas de algunos virus envueltos y las cápsidas de la mayoría de los virus desnudos son eliminados durante el proceso de penetración. Los viriones que penetran parcial o totalmente intactos son desnudados en el interior de la célula, por medio de enzimas hidrolíticas del hospedador, generalmente enzimas lisosómicas.
yessenia severino sanchez mat:89204
ResponderEliminarADHESION
El primer paso en la infección de una célula es la adhesión o fijación a la superficie celular. Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula. Las células que carecen de los receptores apropiados no son susceptibles al virus.
PENETRACIÓN
Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus.
Virus envueltos
(A) Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular.
(B) Entada vía endosomas en la superficie celular.
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
Virus no envueltos o desnudos
Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
EMSAMBLAJE Y MADURACION
Ensamblaje de los componentes de los virus para formar nuevas partículas víricas (proceso también llamado maduración).
Los virus desnudos realizan su ensamblaje y maduración en forma intracelular y adquieren capacidad infectante dentro de la célula. Para su salida requieren de la desintegración celular. Los virus envueltos en el último paso al ensamblaje el virion se une con la membrana celular a la salida de la célula, mediante proteínas que se insertan en la superficie interna y exterior de la membrana. El virion naciente brota atraves de la membrana. El ensamblaje y maduración con la expulsión atraves de la superficie celular proporcionan un mecanismo más eficaz de salida, ya que no depende de la desintegración de la célula infectada.
SALIDA
Liberación de los nuevos virus de la célula hospedadora, que ya pueden infectar otras células. Pueden producirse con o sin rotura de la célula, dependiendo de los virus. Los virus desnudos salen por lisis celular lo contrario de los envueltos que salen por gemación.
Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada
ResponderEliminarLa única característica de la multiplicación viral consiste en que, poco después de su interacción con una célula huésped, el virión infectante se desintegra y pierde la infecciosidad que se puede medir. Esta fase del ciclo celular se llama periodo de eclipse; su duración varía dependiendo del virus y de la célula huésped en particular, y termina con la formación de las primeras partículas virales descendientes infecciosas. El periodo de eclipse es, en realidad, de intensa actividad sintética, puesto que la célula se redirige hacia la satisfacción de las necesidades del "pirata" viral. En algunos casos, tan pronto como el ácido nucleico viral penetra a la célula huésped, el metabolismo celular es recanalizado exclusivamente hacia la síntesis de nuevas partículas virales. En otros casos, los procesos metabólicos de la célula huésped no se alteran significativamente, aunque la célula sintetiza proteínas virales y ácidos nucleicos.
Los virus han desarrollado diversas estrategias para lograr la multiplicación en las células huéspedes parasitadas por ellos. Aunque los detalles varían de un grupo a otro, el patrón general de los ciclos de replicación es semejante.
Virus no envueltos o desnudos
Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
-La primera etapa de la infección viral es la interacción de un virión con un sitio receptor específica de la superficie celular. Las moléculas receptoras difieren para los distintos virus, y son proteínas en algunos casos (por ejemplo, picornavirus) y oligosacáridos en otros (por ejemplo, ortovirus y paramixovirus). La presencia o la ausencia de receptores desempeña una función determinante de primer orden en el tropismo celular y la patogenia viral; por ejemplo, el poliovirus es capaz de fijarse sólo en las células del sistema nervioso central y las vías digestivas de los primates. Se cree que la fijación en receptores refleja homologías configurativas fortuitas entre la estructura de la superficie del virión y un componente de la superficie celular. Por ejemplo, el virus de la inmunodeficiencia humana (HIV) se une al receptor CD4 en las células del sistema inmunitario y se cree que el virus de la rabia interactúa con los receptores de acetilcolina y que el virus de Epstein-Barr reconoce al receptor para el tercer componente del complemento en las células B. Es probable que cada célula susceptible posea por lo menos 100,000 sitios receptores para un virus dado
-mediante Después de la fijación, la partícula viral entra en la célula. Esta etapa se conoce como penetración, viropexia o fagocitosis. En algunos sistemas, se realiza por endocitosis mediada por receptores, con captación de las partículas virales ingeridas dentro de endosomas. Están menos claros los detalles de la penetración en otros sistemas. Ocurre perdida de la cubierta de manera concomitante con la penetración o poco después de la misma. La pérdida de la cubierta, o descubrimiento, es la separación física del ácido nucleico viral (o, en algunos casos, de las nucleocápsides inteARNs) a partir de los componentes estructurales externos del virión. En este momento se pierde la infecciosidad del virus original.
Los virus son los únicos agentes infecciosos en los que la disolución del agente infectante es una etapa obligatoria de la vía de la replicación